Смена генерација
Смена генерација, позната и као смена фаза и метагенеза, тип је животног циклуса који се јавља код неких биљака и алги у групама Archaeplastida и Heterokontophyta које имају изражене хаплоидне полне и диплоидне бесполне фазе. У тим групама, вишећелијски гаметофит, који има хаплоидан (n) број хромозома, смењује се с вишећелијским спорофитом, који је са диплоидним 2n бројем хромозома, сачињеног од „n” парова. Зрели спорофит ствара споре, путем мејозе, у процесу којим се број хромозома смањује на пола, из 2n на n.[1][2]

Хаплоидне споре клијају и израсту у хаплоидни гаметофит. У зрелости, гаметофит ствара гамете путем митозе, која не мења број хромосома. Два гамета (који потичу из различитих организама исте врсте или из истог организма) оплодњом стварају зигот, који се развија у диплоидни спорофит. Овај циклус, од гаметофита до гаметофита (или подједнако од спорофита до спорофита), начин је на који се све копнене биљке и многе алге подвргавају полној репродукцији.
Однос спорофита и гаметофита варира између различитих група биљака. У оним алгама које имају наизменичне генерације, спорофити и гаметофити су засебни – независни организми који могу али не морају имати сличан изглед. Код маховина, јетрењача и Anthocerotophyta, спорофит је мање развијен од гаметофита и у великој мери зависи од њега. Иако спорофити маховине могу да врше фотосинтезу, потребна је додатна фотосинтеза из гаметофита како би се одржао раст и развој спора и зависи од њега за опскрбу водом, минералним храњивим материјама и азотом. Насупрот томе, у свим модерним васкуларним биљкама гаметофит је мање развијен од спорофита, иако су њихови девонски преци имали гаметофите и спорофите приближно једнаке сложености. Код папрати, гаметофит је мали и спљоштен аутотрофни проталус на којем је млади спорофит краткотрајно зависан у својој прехрани. У цветница је смањење гаметофита много екстремније; састоји се од само неколико ћелија које у потпуности расту унутар спорофита.
Животиње се другачије развијају. Они директно производе хаплоидне гамете. Не настају хаплоидне споре које би се могле поделити, тако да немају хаплоидни гаметофит који се наизменично формира с диплоидним спорофитом. (Неки инсекти имају посебан систем одређивања пола, при чему се хаплоидни мужјаци производе од неоплођених јајашаца; међутим, женке су диплоидне). Животни циклуси биљака и алги са наизменичним хаплоидним и диплоидним вишећелијским стадијима називају се „диплохаплонтски” (еквивалентни појмови хаплодиплонтски, диплобионтски или дибионтски такође су у употреби). Животни циклуси, као што су код животиња у којима постоји само диплоидна вишећелијска фаза, називају се „диплонтски” (диплонт). Животни циклуси у којима постоји само хаплоидна вишећелијска фаза називају се „хаплонтски” (хаплонт).
Дефиниција уреди
Смена генерација дефинише се као смена вишећелијских диплоидних и хаплоидних облика у животном циклусу организма, без обзира да ли су ти облици слободни и не.[3] Код неких врста, као што је алга Ulva lactuca, диплоидни и хаплоидни облици заиста су независни организми који живе слободно, углавном једнаки по изгледу и стога се каже да су изоморфни. Слободно пливајући хаплоидни гамети творе диплоидни зигот који клија у вишећелијски диплоидни спорофит. Спорофит ствара слободно пливајуће хаплоидне споре које клијају у хаплоидне гаметофите.[4]
Међутим, у неким другим групама, или спорофит или гаметофит је врло смањен и није способан за слободно живљење. На пример, код свих маховина генерација гаметофита је доминантна, а спорофит зависи од ње. Супротно томе, код свих модерних васкуларних копнених биљака гаметофити су јако редуковани, иако фосилни докази упућују на то да су изведени од изоморфних предака. Код биљка семењача, женски гаметофит се потпуно развија унутар спорофита који га штити и негује, те ембриона спорофита који производи. Зрно полена, које је мушки гаметофити, своди се на само неколико ћелија (по само три у многим случајевима). Овдје је појам две генерације мање очит; као што кажу Батеман & Димицхеле: „(с)порофити и гаметофити делотворно функционишу као јединствен организам”.[5] Алтернативни термин 'смена фаза' би вероватно био подеснији.[6]
Историја уреди
Расправе о смени генерација у раном двадесетом веку могу бити збуњујуће, јер постоје различити начини класификовања „генерација” (сексуални насупрот асексуалном, гаметофити насупрот спорофита, хаплоидни – диплоидни итд.).[7] У почетку су Шамисо и Стенструп описали сукцесију различито организованих генерација (сексуалних и асексуалних) код животиња као „наизменична измена генерација”, проучавајући развој животиња плашташа, жарњака и метиља. Овај феномен је такође познат као хетерогамија. Сада се појам „смена генерација” готово искључиво повезује са животним циклусима биљака, тачније са изменом хаплоидних гаметофита и диплоидних спорофита.[7]
Вилхелм Хофмејстер показао је морфолошку смену генерација код биљска,[2] између генерације која носи споре (спорофит) и генерације која подноси гамете (гаметофит).[8][9][10] У то се време отворила расправа која се усредоточила на порекло асексуалне генерације копнених биљака (тј. спорофита) и конвенцијски је окарактеризирана као сукоб између теорија: антитеза (Челаковски 1874) и хомологије (Прингшајм, 1876) смена генерација.[7] Према некима, Челаковски је сковао речи спорофит и гаметофит.
Едуард Страсбургер (1874) открио је смену генерација између диплоидне и хаплоидне фазе једра,[7] звану цитолошка смена једрених фаза.[11] Иако се најчешће подударају морфолошка смена и смена једарних фаза понекад су независне једна од друге, нпр. код многих црвених алги, иста нуклеусна фаза може одговарати двема различитим морфолошким генерацијама. Код неких папрати које су изгубиле сексуалну репродукцију, у нуклеусној фази нема промене, али смена генерација се одржава.[12]
Смена генерација код биљака уреди
Основни елементи уреди
Горњи дијаграм приказује основне елементе смене генерација код биљака. Многе варијације које су пронађене у различитим групама биљака описане су кориштењем ових концепата касније у чланку. Полазећи с десне стране дијаграма, укључени су следећи процеси:
- Два једноћелијска хаплоидна гамета, од којих сваки садржи неупарене хромозоме n, спајају се тако да формирају једноћелијски диплоидни зигот, која сада садржи n парова хромозома, односно укупно 2n.
- Једноћелијски диплоидни зигот клија, делећи се нормалним процесом (митозе), који одржава број хромозома на 2n. Резултат је мултићелијски диплоидни организам, назван спорофит (јер у зрелости производи споре).
- Када достигне зрелост, спорофит ствара једну или више спорангија, које су органи који производе диплоидне матичне ћелије споре (спороцити). Оне се деле посебним поступком (мејоза) који смањује број хромозома за половину. То у почетку резултира у четири једноћелијске хаплоидне споре, а свака садржи n неупарених хромозома.
- Једноћелијска хаплоидна спора клија, деливши се нормалним процесом (митозом), који одржава број хромозома на n. Резултат је мултићелијски хаплоидни организам, назван гаметофит (јер у зрелости ствара гамете).
- Када достигне зрелост, гаметофит ствара једну или више гаметангија, које су органи за производњу хаплоидних гамета. Барем једна врста гамета поседује неки механизам за достизање другог гамета да би се с њим спојила.
„Смена генерација” у животном циклусу је тако између диплоидне (2n) генерације спорофита и хаплоидне (n) генерације гаметофита.

Ситуација је прилично различита од оне код животиња, где је темељни процес да диплоидна (2n) јединка директно ствара хаплоидне (n) гамете мејозом. Споре (тј. хаплоидне ћелије које су у стању да се подвргну митози) не настају, тако да нема ни асексуалне мултићелијске генерације која се смењује са сексуалном вишећелијском генерацијом. (Неки инсекти – као пчеле, на пример, имају хаплоидне мужјаке који се развијају из неоплођених јајашаца, док су све женке диплоидне.)
Варијације уреди
Дијаграм који је горе приказан добар је приказ животног циклуса неких вишећелијских алги (нпр. род Cladophora), који има спорофите и гаметофите готово идентичног изгледа и који немају различите врсте спора или гамета.[13] Међутим, постоји много могућих варијација темељних елемената животног циклуса који имају смену генерација. Свака варијација може се појавити одвојено или у комбинацији, што резултира чудесном разноликошћу животних циклуса. Изрази које користе ботаничари у описивању ових животних циклуса могу бити подједнако збуњујући. Као што кажу Батеман и Димичел „[...] смена генерација је постала терминолошка мора; често један појам представља неколико појмова или је један појам представљен с више појмова”.[14]


Могуће варијације су:
- Релативни значај спорофита и гаметофита
- Једнаке (хомоморфија или изоморфија).
Кончасте алге из рода Cladophora, које се углавном налазе у свежем водама, имају диплоидне спорофите и хаплоидне гаметофите који су споља неразлучиви. Еволуција смене генерација сугерира да су то чинили њихови преци.[15] Ни једна данашња копнена биљка нема подједнако доминантне спорофите и гаметофите, мада неке теорије еволуције смене генерација сугеришу да су то чиниле и копнене предачке биљке. - Неједнако (хетероморфија или анизоморфија)
- Доминантни гаметофит (гаметофитне).
Код јетрењача, маховина и папрати, доминантни облик је хаплоидни гаметофит. Диплоидни спорофит није способан за самосталан опстанак, растући претежно од родитељског гаметофита, а хлорофил добија тек када сазри.[16] - Доминантни спорофит (спорофитне).
Код папрати су спорофит и гаметофит способни за самосталан живот, али доминантан облик је диплоидни спорофит. Хаплоидни гаметофит је много мањи и једноставније грађе. Код биљки семењача, гаметофит се још више смањује (најмањи је, са само три ћелије), сву своју храну добија од спорофита. Екстремно смањење величине гаметофита и његово задржавање унутар спорофита значи да када се примењује на семенским биљкама, израз „смена генерација” помало доводи у заблуду: „[с]порофит и гаметофит ефикасно функционишу као јединствен организам”.[5] Неки аутори преферирају термин „смена фаза”.[6]
- Доминантни гаметофит (гаметофитне).
- Једнаке (хомоморфија или изоморфија).

- Диференцијација гамета
- Оба гамета су једнака (изогамија).
Као и друге врсте рода Cladophora, C. callicoma има бичасте гамете који су идентичног изгледа и способности кретања.[15] - Гамети различите величине (анизогамија).
- Оба гамета сличне покретљивости.
Врсте морске салате Ulva имају гамете са по два бича за кретање. Они су, међутим, различите величине: већи су женски, а мањи мушки.[17] - Један велики и сесилан, други мален и покретан (оогамија). Већи седећи мегагамети су јајашца (јајне ћелије), а мањи покретни микрогамети су сперматозоиди. Степен покретљивости спермија може бити врло ограничен (као у случају цветница), али сви се могу кретати према седећим јајима. Када се (као што је готово увек случај) стварају сперматозоиди и јајашца у различитим врстама гаметангија, оне које стварају сперму називају се антеридије, а оне које стварају јајашца архегонија (архегонијум).
- Антеридијум и архегонијум јављају се на истом гаметофиту, званом једнодомни. (Многи извори, укључујући оне који се баве маховинама, користе израз „моноецијски” за ову ситуацију, а „диецијски” за супротно.[18][19] Овде се „моноецијски” и „диецијски” употребљавају само за спорофите.)
Јетрењача Pellia epiphylla има гаметофит као доминантну генерацију. Она је моноецијска: мала антеридија која производи црвенкасту сперму распршена је по средини, док архегонија (која производи јаја) расте ближе врховима подела биљке.[20] - Антеридијум и архегонијум појављују се на различитим гаметофитима, који су диецијски.
Маховина Mnium hornum има гаметофит као доминантну генерацију. Она је диецијска: мушке биљке стварају само антеридије у вршним розетама, а женске само архегоније у облику стабљичастих капсула.[21] Семе биљних гаметофита је такође двослојно. Међутим, родитељски спорофит може бити једноличан, производећи мушке и женске гаметофите или диецијски производећи гаметофите само једног пола. Биљке семењаче имају гаметофит изузетно смањене величине; архегонијум се састоји од само малог броја ћелија, а цео мушки гаметофит може бити представљен са само две ћелије.[22]
- Антеридијум и архегонијум јављају се на истом гаметофиту, званом једнодомни. (Многи извори, укључујући оне који се баве маховинама, користе израз „моноецијски” за ову ситуацију, а „диецијски” за супротно.[18][19] Овде се „моноецијски” и „диецијски” употребљавају само за спорофите.)
- Оба гамета сличне покретљивости.
- Оба гамета су једнака (изогамија).
- Диференцијација спора
- Све споре исте величине (хомоспорија или изоспорија)
коњски репови (врсте рода Equisetum) имају споре које су све исте величине. - Две споре различите величине (хетероспорија или анизоспорија): веће мегаспоре и мање микроспоре. Кад се две врсте спора производе у различитим врстама спорангија, оне се називају мегаспорангије и микроспорангије. Мегаспора се често (али не увек) развија на штету остале три ћелије проистекле из мејозе, које абортирају.
Гаметофити семењача такође су диецијки. Међутим, родитељски спорофит може бити једноличан, производећи мушке и женске гаметофите или дводоми, производећи гаметофите само једног пола. Њихови аметофити су изузетно смањене величине; архегонијум се састоји од малог броја ћелија, а цео мушки гаметофит може бити представљен са само две ћелије.- Мегаспорангија и микроспорангија се јављају на истом спорофиту, који се тада зове моноецијски.
Већина цветница спада у ову категорију. Тако цвет љиљана садржи шест стабљика (микроспорангије) које стварају микроспоре; оне се развијају у поленово зрно (микрогаметофит) и три спојена карпела који стварају интегриране мегаспорангије (овуле) од којих сваки ствара мегаспору. Она се развија у мегаспорангију, да би произвела мегагаметофит. У другим биљкама, као што је лешник, неки цветови имају само стабљике, друге само прашнике, али иста биљка (тј. порофит) има обе врсте цвета и тако је моноецијска.
Цветови европске божиковине, дводоме врсте: мушка изнад, женска доле (лишће је одрезано да би се јасније приказали цветови) - Мегаспорангија и микроспорангија на различитим спорофитима који су, дакле диецијски (дводоми).
Појединачна стабла европске боћиковине (Ilex aquifolium) производе или 'мушке' цветове који имају само функционалне стабљике (микроспорангије), производећи микроспоре које се развијају у поленово зрно (микрогаметофити) или 'женски' цветови које има само функционалну карпелу која производи интегриране мегаспорангије (овуле); оне садрже мегаспоре које се развија у вишећелијски мегагаметофит.
- Мегаспорангија и микроспорангија се јављају на истом спорофиту, који се тада зове моноецијски.
- Све споре исте величине (хомоспорија или изоспорија)

Између тих варијација постоје неке корелације, али оне су управо то, корелације, а не и апсолутни односи. На пример, код цветница микроспоре на крају производе микрогамете (сперму), а мегаспоре мегагамете (јајашца). Међутим, код папрати и њихових сродника постоје групе са недиференцираним спорама, али диференцираним гаметофитима. На пример, папрат Ceratopteris talictrioidi има споре само једне врсте, које се непрекидно разликују по величини. Мање споре имају тенденцију клијања у гаметофите који производе само антеридијум, који производи сперму.
Комплексни животни циклус уреди

Дијаграм приказује смену генерација код врсте која је хетероморфна, спорофитска, оогаметска, диецијска, хетероспорна и двоприродна. Пример семењаче може бити стаблашица врба (већина врста рода Salix је двополна).[23] Полазећи од средишта дијаграма, укључени процеси су:
- Непомично јаје, садржано у архегонију, стапа се с покретном спермом, ослобођеном антеридијом. Резултирајући зигот је или мушки или женски'.
- Мушки зигот развија се митозом у микроспорофит који у зрелости производи једну или више микроспорангија. Микроспоре се мејозом развијају у микроспорангију.
Код врбе (попут свих семењача) зигот се најпре развија у микроспорофит ембриона, унутар овуле (мегаспорангија затворена у један или више заштитних слојева ткива познатих као интегумент). Сазревањем ове структуре постаје семе. Касније се семе расипа, клија и израсте у зрело дрво. „Мушко” стабло врбе (микроспорофит) производи цветове са само пршницима, од којих су антере микроспорангије. - Микроспоре клијају производећи микрогаметофите; у зрелости настаје једна или више антхеридија. Сперматозоиди се развијају у антхеридији.
Код врбе, микроспоре се не ослобађају од антера (микроспорангија), већ се развијају у поленова зрна (микрогаметофити) унутар ње. Расијава се целокупно зрно полена (нпр. инсектима или ветром) у овуле (мегагаметофит), где се ствара сперма која се креће низ поленову цев до јајне ћелије. - Женски зигот развија се путем митозе у мегаспорофит, који кад сазри, производи једну или више мегаспорангија. Мегаспоре се развијају унутар мегаспорангије; типски, једна од четири споре произведене мејозом развијају се на рачун преостале три, које нестају.
'Женске' врбе (мегаспорофити) производе цветове само са тучковима (модификовано лишће које носи мегаспорангију). - Мегаспоре клијају стварајући мегагаметофите; у зрелости се производи једна или више архегонија. Јаја се развијају у архегонији.
Карпеле врбе производе овуле, мегаспорангије су затворене у интегументе. Унутар сваке овуле митозом се развија мегаспора у мегагаметофит. Архегонијум се развија унутар мегагаметофита и ствара јаје. Читава „гаметофитска генерација” остаје под заштитом спорофита, осим поленових зрнаца (сведена у три ћелије које се налазе у зиду микроспоре).
- Мушки зигот развија се митозом у микроспорофит који у зрелости производи једну или више микроспорангија. Микроспоре се мејозом развијају у микроспорангију.
Животни циклус по групама биљака уреди
Термин „биљке” укључује Archaeplastida, тј. Glaucophyta, црвене и зелене алге и копнене биљке. Смена генерација се јавља у готово свим вишећелијским црвеним и зеленим алгама, код слатководних форми (као што је Cladophora) и морских (као што су оне у роду Ulva). Код већине њих генерације су хомоморфне (изоморфне) и слободно живе. Неке врсте црвених алги имају сложену трофазну смену генерација у којој постоји фаза гаметофита и две различите спорофитне фазе. За додатне информације погледајте црвене алге: репродукција.
Све копнене биљке имају хетероморфну (анизоморфну) смену генерација, у којој се спорофити и гаметофити изразито разликују. Код свих припадника групе Bryophyte, тј. јетрењача, маховина и роговника, генерације гаметофита се најочитије испољавају. Као илустрација се може размотрити једнодомна маховина. Антеридија и архегонија се развијају на зрелој биљци (гаметофит). У присуству воде, бифлагелатни сперматозоиди из антеридије допливају до архегоније и долази до оплодње, што доводи до стварања диплоидног спорофита. Спорофити расту из архегонија. Њихово тело састоји се од дуге стабљике на врху капсуле унутар које ћелије које производе споре пролазе кроз мејозу и формирају хаплоидне споре. Код већине маховина споре се распршују ветром, иако је Splachnum sphaericum ентомофилна, привлачи инсекте како би раширила своје споре. За додатне информације погледајте јетрењаче: животни циклус, маховине: животни циклус, роговници: животни циклус.
-
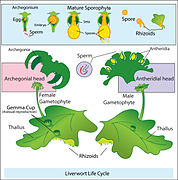 Дијаграм смене генерација код јетрењача
Дијаграм смене генерација код јетрењача -
 Дијаграм животног циклуса маховине
Дијаграм животног циклуса маховине -
 Дијаграм животног циклуса роговника
Дијаграм животног циклуса роговника



Код папрати и њиховим сродницима, укључујући преслицу и коњски реп, уочљивија биљка у пољу је диплоидни спорофит. Хаплоидне споре развијају се у сорусима на наличју и расипају се ветром (или у неким случајевима, плутајући по води). Ако су услови добри, спора ће клијати и израсти у прилично неупадљиво биљно тело звано проталус. Хаплоидни проталус не личи на спорофит, па као такве папрати и њихови сродници имају хетероморфну смену генерација. Проталус је краткотрајан, али има полно размножавање, производећи диплоидни зигот који из проталуса расте као спорофит. За додатне информације погледајте папрат#животни циклус.
-

Дијаграм смене генерација код папрати. -

Гаметофит (проталус) Dicksonia sp. -

Спорофит Dicksonia antarctica. -

Наличје Dicksonia antarctica са сорусима – структурама за стварање спора




У групи Spermatophyte, семењачама, спорофит је доминантна вишећелијска фаза; гаметофити су јако смањени и врло се разликују по морфологији. Читава генерација гаметофита, с изузетком поленових зрна (микрогаметофити), налази се унутар спорофита. Животни циклус биљке цветнице (ангиосперме), врбе, детаљно је описана у претходном одељку. Животни циклус голосеменица је сличан. Међутим, цветнице имају и феномен који се назива „двострука оплодња”. Два једра сперме из поленовог зрна (микрогаметофит), а не једно, улазе у архегонијум мегагаметофита; једно се стапа с једром јајашца за формирање зигота, а друго са два друга једра гаметофита, како би се формирао ендосперм, који храни ембрион који се развија. За додатне информације погледајте двострука оплодња.
Еволуција доминантности диплоидне фазе уреди
Предложено је да је основа за настанак диплоидне фазе животног циклуса (спорофита) као доминантне фазе (нпр. код васкуларних биљака) то што диплоидија омогућава маскирање експресије штетних мутација путем генетичке комплементације.[24][25]
Према томе, ако је један од родитељских генома у диплоидним ћелијама садржавао мутацију која доводи до оштећења у једном или више генских производа, ти би недостаци могли бити надокнађени другим родитељским геномом (који ипак може имати своје властите мане у другим генима). Како је диплоидна фаза постајала превладавајућа, учинак маскирања вероватно је омогућио да се величина генома, а тиме и садржај информација, повећа без ограничења на то да се мора побољшати прецизност репликације ДНК. Могућност повећања садржаја информација по ниској цени била је повољна, јер је омогућила шифрирање нових прилагођавања. Ово гледиште је оспоравано, а докази показују да селекција није ефикаснија у хаплоидној него у диплоидној фази животног циклуса маховина и ангиосперми.[26]
-
 Животни циклус ангиосперми
Животни циклус ангиосперми -
 Врх тулипановог прашника са поленом (микрогаметофити)
Врх тулипановог прашника са поленом (микрогаметофити) -
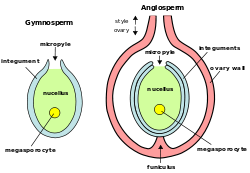 Биљне овуле (мегагаметофити): Овуле голосемењача (лево) и ангиосперми
Биљне овуле (мегагаметофити): Овуле голосемењача (лево) и ангиосперми -
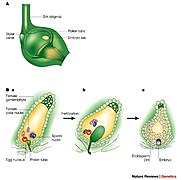 Двострука оплодња
Двострука оплодња




Слични процеси код осталих организама уреди
Rhizaria уреди
Неки организми, који су сада класификовани у кладус Rhizaria и биљке у смислу какав се овде користи не показују смену генерација. Већина Foraminifera пролази кроз хетероморфну смену генерација између хаплоидних гамонт и диплоидних агамонт облика. Једноћелијски хаплоидни организам је обично много већи од диплоидног организма.
Гљиве уреди
Мицелијуми гљива су типски хаплоиди. Кад се сусретну два упарујућа мицелијума различитих врста, стварају две мултиједарне ћелије у облику куглице, које се спајају преко „моста за парење”. Једра прелазе из једног мицелијума у други, формирајући хетерокарион (што значи „различита једра”). Овај процес се назива плазмогамија. Стварно стапање у диплоидна једра назива се кариогамија и не може се догодити док се не формирају спорангије. Карогамија производи диплоидни зигот, што је краткотрајни спорофит који убрзо пролази кроз мејозу да би формирао хаплоидне споре. Кад споре клијају, развијају се у нов мицелијум.
Слузаве плесни уреди
Животни циклус слузавих плесни је врло сличан животињском. Хаплоидне споре клијају у творевине с гомилом ћелија или myxamoebae. Оне се спајају у процесу који се назива плазмогамија и кариогамија да би се формирао диплоидни зигот. Жигот се развија у плазмодијум, а кад овај сазри производи од једног до много плодних тела која садрже хаплоидне споре, зависно од врсте.
Животиње уреди
Смена између вишећелијске диплоиде и вишећелијске хаплоидне генерације се никад не сусреће код животиња. Код неких животиња постоји смена између фаза партеногенезе и полне репродукције (хетерогамија). Обе фазе су диплоидне. То се понекад назива „смена генерација”, мада је сасвим другачија. Код неких других животиња, као што су инсекти из реда Hymenoptera (где спада и пчела), мужјаци су хаплоидни, а женке диплоидне, и то је увек случај, уместо да постоји смена између различитих генерација.
Види још уреди
Референце уреди
- ^ Керп, Х.; Треwин, Н.Х. & Хасс, Х. (2003), „Неw гаметопхyтес фром тхе Лоwер Девониан Рхyние Цхерт”, Трансацтионс оф тхе Роyал Социетy оф Единбургх: Еартх Сциенцес, 94 (4): 411—428, дои:10.1017/С026359330000078X
- ^ а б Сведелиус, Нилс (1927), „Алтернатион оф Генератионс ин Релатион то Редуцтион Дивисион”, Ботаницал Газетте, 83 (4): 362—384, ЈСТОР 2470766, дои:10.1086/333745
- ^ Таyлор, Керп & Хасс 2005
- ^ "„Плант Сциенце 4 У”. Приступљено 5. 7. 2016.
- ^ а б Батеман & Димицхеле 1994, стр. 403
- ^ а б Стеwарт & Ротхwелл 1993
- ^ а б в г Хаиг, Давид (2008), „Хомологоус версус антитхетиц алтернатион оф генератионс анд тхе оригин оф споропхyтес” (ПДФ), Тхе Ботаницал Ревиеw, 74 (3): 395—418, дои:10.1007/с12229-008-9012-x, Приступљено 17. 8. 2014
- ^ Хофмеистер, W. (1851), Верглеицхенде Унтерсуцхунген дер Кеимунг, Ентфалтунг унд Фруцхтбилдилдиунг хöхерер Крyптогамен (Моосе, Фарне, Еqуисетацеен, Рхизоцарпеен унд Лyцоподиацеен) унд дер Саменбилдунг дер Цониферен (на језику: Герман), Леипзиг: Ф. Хофмеистер, Приступљено 17. 8. 2014
- ^ Цурреy, Фредерицк (1862), Он тхе герминатион, девелопмент, анд фруцтифицатион оф тхе хигхер Црyптогамиа, анд он тхе фруцтифицатион оф тхе Цониферæ, Лондон: Роберт Хардwицке, Приступљено 17. 8. 2014
- ^ Фелдманн, Ј. & Фелдманн, Г. (1942), „Рецхерцхес сур лес Боннемаисониацéес ет леур алтернанце де генератионс” (ПДФ), Анн. Сци. Натл. Бот., Сер. 11 (на језику: Френцх), 3: 75—175, Архивирано из оригинала (ПДФ) 19. 08. 2014. г., Приступљено 08. 10. 2019, п. 157
- ^ Фелдманн, Ј. (1972), „Лес проблèмес ацтуелс де л'алтернанце де гéнератионс цхез лес Алгуес”, Буллетин де ла Социéтé Ботаниqуе де Франце (на језику: Френцх), 119: 7—38, дои:10.1080/00378941.1972.10839073
- ^ Сцхопфер, П.; Мохр, Х. (1995). „Пхyсиологy оф Девелопмент”. Плант пхyсиологy. Берлин: Спрингер. стр. 288—291. ИСБН 978-3-540-58016-4.
- ^ Гуирy & Гуирy 2008
- ^ Батеман & Димицхеле 1994, стр. 347
- ^ а б Схyам 1980
- ^ Wатсон 1981, стр. 2
- ^ Кирбy 2001
- ^ Wатсон 1981, стр. 33
- ^ Белл & Хемслеy 2000, стр. 104
- ^ Wатсон 1981, стр. 425–6
- ^ Wатсон 1981, стр. 287–8
- ^ Спорне 1974а, стр. 17–21.
- ^ „Wиллоwс”, Енцyцлопæдиа Британница, XIX (11тх изд.), Неw Yорк: Енцyцлопæдиа Британница, 1911, Приступљено 1. 1. 2011
- ^ Бернстеин, Х.; Бyерс, Г.С. & Мицход, Р.Е. (1981), „Еволутион оф сеxуал репродуцтион: Импортанце оф ДНА репаир, цомплементатион, анд вариатион”, Тхе Америцан Натуралист, 117 (4): 537—549, дои:10.1086/283734
- ^ Мицход, Р.Е. & Гаyлеy, Т.W. (1992), „Маскинг оф мутатионс анд тхе еволутион оф сеx”, Тхе Америцан Натуралист, 139 (4): 706—734, дои:10.1086/285354
- ^ Сзöвéнyи, Пéтер; Рицца, Мариана; Хоцк, Зсóфиа; Схаw, Јонатхан А.; Схимизу, Кентаро К. & Wагнер, Андреас (2013), „Селецтион ис но море еффициент ин хаплоид тхан ин диплоид лифе стагес оф ан ангиосперм анд а мосс”, Молецулар Биологy анд Еволутион, 30 (8): 1929—39, ПМИД 23686659, дои:10.1093/молбев/мст095
Литература уреди
- Барнес, Р.С.К.; Цалоw, П.; Оливе, П.Ј.W.; Голдинг, D.W. & Спицер, Ј.I. (2001), Тхе Инвертебратес: а сyнтхесис, Оxфорд; Малден, МА: Блацкwелл, ИСБН 978-0-632-04761-1
- Батеман, Р.M. & Димицхеле, W.А. (1994), „Хетероспорy – тхе мост итеративе кеy инноватион ин тхе еволутионарy хисторy оф тхе плант кингдом” (ПДФ), Биологицал Ревиеwс оф тхе Цамбридге Пхилосопхицал Социетy, 69 (3): 345—417, дои:10.1111/ј.1469-185x.1994.тб01276.x, Архивирано из оригинала (ПДФ) 15. 04. 2012. г., Приступљено 30. 12. 2010
- Белл, П.Р. & Хемслеy, А.Р. (2000), Греен Плантс: тхеир Оригин анд Диверситy (2нд изд.), Цамбридге, етц.: Цамбридге Университy Пресс, ИСБН 978-0-521-64109-8
- Фостер, А.С. & Гиффорд, Е.M. (1974), Цомпаративе Морпхологy оф Васцулар Плантс (2нд изд.), Сан Францисцо: W.Х. Фрееман, ИСБН 978-0-7167-0712-7
- Гуирy, M.D.; Гуирy, Г.M. (2008), „Цладопхора”, АлгаеБасе, Wорлд-wиде елецтрониц публицатион, Натионал Университy оф Иреланд, Галwаy, Приступљено 21. 7. 2011
- Кирбy, А. (2001), Улва, тхе сеа леттуце, Монтереy Баy Аqуариум Ресеарцх Институте, Архивирано из оригинала 16. 5. 2011. г., Приступљено 1. 1. 2011
- Сцотт, Тхомас (1996), Цонцисе Енцyцлопедиа Биологy, Берлин: Wалтер де Груyтер, ИСБН 978-3-11-010661-9
- Схyам, Р. (1980), „Он тхе лифе-цyцле, цyтологy анд таxономy оф Цладопхора цаллицома фром Индиа”, Америцан Јоурнал оф Ботанy, 67 (5): 619—24, ЈСТОР 2442655, дои:10.2307/2442655
- Спорне, К.Р. (1974а), Тхе Морпхологy оф Ангиоспермс, Лондон: Хутцхинсон, ИСБН 978-0-09-120611-6
- Спорне, К.Р. (1974б), Тхе Морпхологy оф Гyмноспермс (2нд изд.), Лондон: Хутцхинсон, ИСБН 978-0-09-077152-3
- Стеwарт, W.Н. & Ротхwелл, Г.W. (1993), Палеоботанy анд тхе Еволутион оф Плантс (2нд изд.), Цамбридге, УК: Цамбридге Университy Пресс, ИСБН 978-0-521-38294-6
- Wатсон, Е.V. (1981), Бритисх Моссес анд Ливерwортс (3рд изд.), Цамбридге, УК: Цамбридге Университy Пресс, ИСБН 978-0-521-28536-0
- Таyлор, Т.Н.; Керп, Х. & Хасс, Х. (2005), „Лифе хисторy биологy оф еарлy ланд плантс: Деципхеринг тхе гаметопхyте пхасе”, Процеедингс оф тхе Натионал Ацадемy оф Сциенцес оф тхе Унитед Статес оф Америца, 102 (16): 5892—5897, ПМЦ 556298
 , ПМИД 15809414, дои:10.1073/пнас.0501985102
, ПМИД 15809414, дои:10.1073/пнас.0501985102
Спољашње везе уреди
- Generationswechsel. Spektrum, Heidelberg 1999.
- Sexualität bei Ginkgo