Mlečna žlezda
Mlečna žlezda je endokrina žlezda kod ljudi i drugih sisara koja proizvodi mleko za ishranu mladog potomstva. Sisari su svoje ime dobili po latinskoj reči mamma, „dojka“. Mlečne žlezde su raspoređene u organe kao što su dojke kod primata (na primer, ljudi i šimpanze), vime kod preživara (na primer krave, koze i jeleni) i kod još drugih životinja (na primer, pasa i mačke). Galaktoreja, povremena proizvodnja mleka u žlezdama, može se javiti kod bilo kog sisara, ali kod većine sisara, laktacija, proizvodnja dovoljno mlijeka za njegu, javlja se samo kod fenotipskih ženki koje su gestovale poslednjih meseci ili godina. Usmeravaju ga hormonalne smjernice polnih steroida. Kod nekoliko vrsta sisara može doći do muške laktacije. Kod ljudi se muška laktacija može dogoditi samo pod određenim okolnostima.
| Nazivi i oznake | |
|---|---|
| TA98 | A16.0.02.006 |
| TA2 | 7099 |
| FMA | 60088 |
| Anatomska terminologija | |
Sisari su podjeljeni u 3 grupe: prototerijanci, metaterijanci i euterije. U slučaju prototeraca, i muškarci i žene imaju funkcionalne mlečne žlezde, ali njihove mlečne žlezde su bez bradavica. Ove mlečne žlezde su modifikovane lojne žlezde. Što se tiče metaterijanaca i euterija, samo žene imaju funkcionalne mlečne žlezde. Njihove mlečne žlezde mogu se nazvati dojkama ili vimenom. U slučaju dojki, svaka mlečna žlezda ima svoju bradavicu. U slučaju vimena, parovi mlečnih žlezda čine jednu masu, a na nju visi više od jedne bradavice. Na primer, krave i bivoli imaju po jedno vime sa četiri sise, dok ovce i koze imaju po dve sise koji vire iz vimena. Ove mlečne žlezde su modifikovane znojne žlezde.
Struktura
urediOsnovne komponente zrele mlečne žlezde su alveole (velike nekoliko milimetara), koje su obložene kuboidnim ćelijama koje luče mleko i okružene mioepitelnim ćelijama. Ove alveole se spajaju u grupe poznate kao lobuli. Svaka lobula ima mlečni kanal koji se odvodi u otvore na bradavici. Mioepitelne ćelije se kontrahuju pod stimulacijom oksitocina, izlučujući mleko koje luče alveolarne jedinice u lumen lobula prema bradavici. Kako dojenče počinje sisati, dolazi do „refleksa puštanja“ posredovanog oksitocinom, a majčino mleko se izlučuje - a ne usisava - iz žlezde u bebina usta. [4]
Svo tkivo koje luči mleko koje vodi do jednog mlečnog kanala naziva se zajednički „jednostavna mlečna žlezda“; u „složenoj mlečnoj žlezdi“ sve jednostavne mlečne žlezde služe jednoj bradavici. Ljudi obično imaju dve složene mlečne žlezde, po jednu u svakoj dojci, a svaka složena mlečna žlezda sastoji se od 10–20 jednostavnih žlezda. Prisustvo više od dve bradavice poznato je kao politelija, a prisustvo više od dve složene mlečne žlezde kao polimastija.
Održavanje tačne polarizovane morfologije stabla mlečnog kanala zahteva još jednu bitnu komponentu - ekstracelularni matriks mlečnih epitelnih ćelija (ECM), koji zajedno sa adipocitima, fibroblastom, inflamatornim ćelijama i drugim čine mlečnu stromu. [5] ECM mlečnog epitela uglavnom sadrži mioepitelnu bazalnu membranu i vezivno tkivo. Oni ne samo da pomažu u podršci osnovne strukture mlečnih žlezda, već i služe kao komunikacioni most između mlečnih epitela i njihovog lokalnog i globalnog okruženja tokom celog razvoja ovog organa. [6][7]
Histologija
uredi
Mlečna žlezda je specifična vrsta apokrine žlezde specijalizovana za proizvodnju kolostruma tokom porođaja. Mnogi izvori tvrde da su mlečne žlezde modifikovane znojne žlezde. [8][9][10] Neki autori to osporavaju i umesto toga tvrde da su lojne žlezde. [8]
Razvoj
urediMlečne žlezde se razvijaju tokom različitih ciklusa rasta. Postoje u oba pola tokom embrionalne faze, formirajući samo rudimentarno drvo kanala pri rođenju. U ovoj fazi razvoj mlečnih žlezda zavisi od sistemskih (i majčinih) hormona, [5] ali je takođe pod (lokalnom) regulacijom parakrine komunikacije između susednih epitelnih i mezenhimalnih ćelija proteinima povezanim sa paratiroidnim hormonima PTHrP. [11] Ovaj lokalno sekretovani faktor dovodi do niza pozitivnih povratnih informacija iznutra između ove dve vrste ćelija, tako da epitelne ćelije mlečnih pupoljaka mogu da se razmnožavaju i niču u mezenhimalni sloj dok ne dođu do masnog sloja da bi započele prvi krug grananja. [5] Istovremeno, embrionalne mezenhimske ćelije oko epitelnog pupoljka dobijaju faktore sekrecije koje aktivira PTHrP, kao što je BMP4. Ove mezenhimske ćelije mogu se transformisati u gusti, specifični za mlečne žlezde mezenhim, koji se kasnije razvija u vezivno tkivo sa vlaknastim nitima, formirajući krvne sudove i limfni sistem. [12] Bazalna membrana, koja uglavnom sadrži laminin i kolagen, nastala nakon toga diferenciranim mioepitelnim ćelijama, zadržava polaritet ovog primarnog stabla kanala. Ove komponente ekstracelularnog matriksa snažne su odrednice morfogeneze kanala. [13]
Biohemija
urediEstrogen i hormon rasta (GH) su od suštinske važnosti za duktalnu komponentu razvoja mlečnih žlezda i deluju sinergijski da bi to posredovali. [14][15][16][17][18] Ni estrogen ni GH nisu sposobni da indukuju duktalni razvoj jedan bez drugog.[15][16][17][18][15][16][17][18][19] Utvrđeno je da je uloga GH u duktalnom razvoju uglavnom posredovana njegovom indukcijom sekrecije insulinu sličnog faktora rasta (IGF-1), koji se javlja sistemski (uglavnom poreklom iz jetre). [18] Međutim, sam GH takođe deluje nezavisno od IGF-1 da stimuliše razvoj duktala regulacijom ekspresije estrogenskog receptora (ER) u tkivu mlečne žlezde, što je nizvodni efekat aktivacije GHR mlečne žlezde. [18] U svakom slučaju, za razliku od IGF-1, sam GH nije bitan za razvoj mlečnih žlezda, a IGF-1 u sprezi sa estrogenom može da indukuje normalan razvoj mlečnih žlezda bez prisustva GH. [18] Pored IGF-1, prisutni su i drugi parakrini faktori rasta kao što su epidermalni faktor rasta (EGF), transformišući faktor rasta beta (TGF-β), [20] amfiregulin, [21] faktor rasta fibroblasta (FGF) i faktor rasta hepatocita ( HGF) [22] su uključeni u razvoj dojke kao posrednici nizvodno od polnih hormona i GH / IGF-1. [23][24][25]
Tokom embrionalnog razvoja, nivo IGF-1 je nizak i postepeno raste od rođenja do puberteta. [26] U pubertetu nivoi GH i IGF-1 dostižu najviši nivo u životu i estrogen počinje da se luči u velikim količinama kod žena, tada se uglavnom odvija duktalni razvoj. [26] Pod uticajem estrogena raste i stromalno i masno tkivo koje okružuje duktalni sistem mlečnih žlezda. [27] Posle puberteta, nivoi GH i IGF-1 se progresivno smanjuju, što ograničava dalji razvoj do trudnoće, ako se dogodi.[traži se izvor] Tokom trudnoće, progesteron i prolaktin su od suštinske važnosti za posredovanje lobuloalveolarnog razvoja u tkivu mlečne žlezde prepunoj estrogenu, što se dešava u pripremi laktacije i nege. [26].[14][28]
Androgeni kao što je testosteron inhibiraju razvoj mlečnih žlezda posredovanih estrogenom (npr. Smanjenjem lokalne ekspresije ER) aktiviranjem androgenih receptora izraženih u tkivu mlečnih žlezda, [28][29] i zajedno sa relativno niskim nivoima estrogena, uzrok su nedostatak razvijenih mlečnih žlezda kod muškaraca. [30]
Pregled
urediPre rođenja
urediRazvoj mlečnih žlezda karakteriše jedinstveni proces kojim epitel napada invaziju strome. Razvoj mlečne žlezde javlja se uglavnom nakon rođenja. Tokom puberteta, formiranje tubula je povezano sa razgranatom morfogenezom koja uspostavlja osnovnu arborealnu mrežu kanala koji izlaze iz bradavice. [31]
U razvoju, epitel mlečne žlezde neprestano proizvode i održavaju retke epitelne ćelije, nazvane rodovima mlečnih žlezda, za koje se na kraju smatra da potiču od matičnih ćelija nastanjenih u tkivu.
Razvoj embrionalne mlečne žlezde može se podeliti u niz specifičnih faza. U početku se formiranje mlečnih linija koje se protežu između prednjih i zadnjih udova obostrano na svakoj strani srednje linije dešava oko embrionalnog dana 10.5 (E10.5). Druga faza se dešava na E11.5 kada započinje formiranje plakova duž mlečne linije mleka. Ovo će na kraju dovesti do pojave bradavice. Na kraju, treća faza se dešava na E12.5 i uključuje invaginaciju ćelija unutar plakova u mezenhim, što dovodi do anlaže mlečnih žlezda. [32]
Primitivne (matične) ćelije se otkrivaju u embrionu i njihov broj se neprestano povećava tokom razvoja [33]
Rast
urediPostnatalno, mlečni kanali se izdužuju u mlečni jastučić - mlečne žlezde. Zatim, počev od oko četiri nedelje starosti, rast duktalnog mlečnog mleka značajno se povećava sa prodiranjem kanala prema limfnom čvoru. Krajnji pupoljci terminala, visoko proliferativne strukture koje se nalaze na vrhovima upadnih kanala, tokom ove faze se uveliko šire i povećavaju. Ovaj razvojni period karakteriše pojava krajnjih pupoljaka i traje do starosti oko 7-8 nedelja.
U pubertetskom stadijumu mlečni kanali su invadirali do kraja mlečne masne podloge. U ovom trenutku krajnji pupoljci postaju manje proliferativni i smanjuju se u veličini. Bočne grane se formiraju iz primarnih kanala i počinju da ispunjavaju mlečni jastučić mlečne žlezde. Duktalni razvoj opada dolaskom polne zrelosti i prolazi kroz estrousne cikluse (proestrus, estrus, metestrus i diestrus). Kao rezultat estrousnog ciklusa, mlečna žlezda se podvrgava dinamičkim promenama gde ćelije proliferiraju, a zatim regresiraju na uređen način. [34]
Trudnoća
urediTokom trudnoće, duktalni sistemi se brzo šire i formiraju alveolarne strukture unutar grana koje će se koristiti za proizvodnju mleka. Nakon porođaja, laktacija se javlja unutar mlečne žlezde; laktacija uključuje lučenje mleka luminalnim ćelijama u alveolama. Kontrakcija mioepitelnih ćelija koje okružuju alveole dovešće do izbacivanja mleka kroz kanale i u bradavicu za dojenče. Nakon odbijanja dojenčeta, laktacija prestaje i mlečna žlezda se okreće sama sebi, proces koji se naziva involucija. Ovaj proces uključuje kontrolisani kolaps epitelnih ćelija mlečne žlezde gde ćelije kontrolisano započinju apoptozu, vraćajući mlečnu žlezdu nazad u pubertetsko stanje.
Postmenopauza
urediTokom postmenopauze, zbog mnogo nižih nivoa estrogena i zbog nižih nivoa GH i IGF-1, koji se smanjuju sa godinama, tkivo mlečnih žlezda atrofira i mlečne žlezde postaju sve manje.
Klinički značaj
urediTumorigeneza mlečnih žlezda može biti indikovana biohemijski nepravilnim nivoom ekspresije hormona u cirkulaciji ili lokalnih komponenata ECM, [35] ili mehaničkom promenom napetosti mlečne strome. [36] U bilo kojoj od ove dve okolnosti, epitelne ćelije mleka će rasti van kontrole i na kraju rezultirati rakom. Skoro svi slučajevi raka dojke potiču iz lobula ili kanala mlečnih žlezda.
Ostali sisari
urediGeneralno
urediGrudi odrasle žene razlikuju se od većine ostalih sisara koji imaju manje vidljive mlečne žlezde. Broj i položaj mlečnih žlezda varira kod različitih sisara. Isturene grudi i prateće žlezde mogu se nalaziti bilo gde duž dve linije mleka. Generalno, većina sisara razvija mlečne žlezde u parovima duž ovih linija, s brojem koji približno odgovara broju mladih tipično rođenih u isto vreme. Broj sisa varira od 2 (kod većine primata) do 18 (kod svinja). Američki oposum ima 13, jedan od retkih sisara sa neparnim brojem.[37][38]Sledeća tabela navodi broj i položaj sisara i žlezda nađenih kod sisara:
| Vrste[39] | Prednji (torakalno) |
Srednji (abnominalno) |
Iza (ingvinalno) |
Ukupno |
|---|---|---|---|---|
| Koza, Ovca, Konj Morsko prase |
0 | 0 | 2 | 2 |
| Govedo | 0 | 0 | 4 | 4 |
| Mačka | 2 | 2 | 4 | 8 |
| Pas[40] | 4 | 2 | 2 ili 4 | 8 ili 10 |
| Miš | 6 | 0 | 4 | 10 |
| Pacov | 6 | 2 | 4 | 12 |
| Svinja | 6 | 6 | 6 | 18 |
| Surlaši, primati | 2 | 0 | 0 | 2 |
| Američki oposum[37][38] | 0 | 0 | 13 | 13 |
| Južni crvenoboki oposum[41] | 0 | 0 | 25 ili 27 | 25 ili 27 |
Muški sisari obično imaju rudimentarne mlečne žlezde i bradavice, uz nekoliko izuzetaka: muški miševi nemaju bradavice, [42] muški torbari nemaju mlečne žlezde, [43] a muški konji nemaju bradavice i mlečne žlezde. Muška laktacija se retko javlja kod nekih vrsta. [44]
Mlečne žlezde su prave fabrike proteina, [45] a nekoliko laboratorija je izgradilo transgene životinje, uglavnom koze i krave, kako bi proizvodile proteine za farmaceutsku upotrebu. Kompleksni glikoproteini, kao što su monoklonska antitela ili antitrombin, ne mogu da proizvedu genetski modifikovane bakterije, a proizvodnja kod živih sisara je mnogo jeftinija od upotrebe ćelijskih kultura sisara.
Evolucija
urediPostoji mnogo teorija o tome kako su mlečne žlezde evoluirale. Na primer, smatra se da je mlečna žlezda transformisana znojna žlezda, bliže povezana sa apokrinim znojnim žlezdama. [46] Budući da mlečne žlezde ne fosilizuju dobro, teško je podržati takve teorije fosilnim dokazima. Mnoge trenutne teorije zasnivaju se na poređenju između linija živih sisara - monotrema, torbarskih životinja i euterija. Jedna teorija predlaže da su mlečne žlezde evoluirale iz žlezda koje su korišćene za održavanje vlažnih jajašaca ranih sisara [47][48] i bez zaraze [49][50] (monotremi još uvek polažu jaja). Druge teorije sugerišu da su rani sekret direktno koristili izleženi mladi [51] ili da su ga mladi koristili da bi im pomogao da se orijentišu prema majkama. [52]
Smatra se da se laktacija razvila mnogo pre evolucije mlečne žlezde i sisara.
Dodatne slike
uredi-
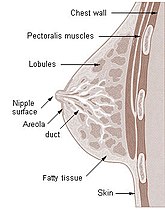 Presek dojke žene
Presek dojke žene -

-

-

-

-

-








Vidi još
urediReference
uredi- ^ Gray, Henry (1918). Anatomy of the Human Body.
- ^ a b v Macéa, José Rafael; Fregnani, José Humberto Tavares Guerreiro (1. 12. 2006). „Anatomy of the Thoracic Wall, Axilla and Breast” (PDF). International Journal of Morphology. 24 (4). doi:10.4067/S0717-95022006000500030
 .
.
- ^ Lawrence, Ruth A.; Lawrence, Robert M. (2010-09-30). Breastfeeding: A Guide for the Medical Profession (7th izd.). Maryland Heights, Maryland: Mosby/Elsevier. str. 54. ISBN 9781437735901.
- ^ Newton, Michael; Newton, Niles Rumely (decembar 1948). „The let-down reflex in human lactation”. The Journal of Pediatrics. 33 (6): 698—704. doi:10.1016/S0022-3476(48)80075-2.
- ^ a b v Watson, C. J.; Khaled, W. T. (2008). „Mammary development in the embryo and adult: A journey of morphogenesis and commitment”. Development. 135 (6): 995—1003. PMID 18296651. doi:10.1242/dev.005439 .
- ^ Wiseman, B. S.; Werb, Z. (2002). „Stromal Effects on Mammary Gland Development and Breast Cancer”. Science. 296 (5570): 1046—1049. Bibcode:2002Sci...296.1046W. PMC 2788989 . PMID 12004111. doi:10.1126/science.1067431.
- ^ Pavlovich, A. L.; Manivannan, S.; Nelson, C. M. (2010). „Adipose Stroma Induces Branching Morphogenesis of Engineered Epithelial Tubules”. Tissue Engineering Part A. 16 (12): 3719—3726. PMC 2991209 . PMID 20649458. doi:10.1089/ten.TEA.2009.0836.
- ^ a b Ackerman (2005) ch.1 „Apocrine Units”. Arhivirano iz originala 21. 04. 2011. g.
- ^ Moore (2010) ch.1 Thorax, p. 99
- ^ Krstic, Radivoj V. (18. 3. 2004). Human Microscopic Anatomy: An Atlas for Students of Medicine and Biology. Springer. str. 466. ISBN 9783540536666.
- ^ Wysolmerski, J. J.; Philbrick, W. M.; Dunbar, M. E.; Lanske, B.; Kronenberg, H.; Broadus, A. E. (1998). „Rescue of the parathyroid hormone-related protein knockout mouse demonstrates that parathyroid hormone-related protein is essential for mammary gland development”. Development. 125 (7): 1285—1294. PMID 9477327. doi:10.1242/dev.125.7.1285.
- ^ Hens, J. R.; Wysolmerski, J. J. (2005). „Key stages of mammary gland development: Molecular mechanisms involved in the formation of the embryonic mammary gland”. Breast Cancer Research. 7 (5): 220—224. PMC 1242158 . PMID 16168142. doi:10.1186/bcr1306.
- ^ Montévil, Maël; Speroni, Lucia; Sonnenschein, Carlos; Soto, Ana M. (2016-10-01). „Modeling mammary organogenesis from biological first principles: Cells and their physical constraints”. Progress in Biophysics and Molecular Biology. From the Century of the Genome to the Century of the Organism: New Theoretical Approaches. 122 (1): 58—69. PMC 5563449 . PMID 27544910. doi:10.1016/j.pbiomolbio.2016.08.004.
- ^ a b Brisken; Malley (2. 12. 2010). „Hormone Action in the Mammary Gland”. Cold Spring Harbor Perspectives in Biology. 2 (12): a003178. PMC 2982168 . PMID 20739412. doi:10.1101/cshperspect.a003178.
- ^ a b v Kleinberg DL (1998). „Role of IGF-I in normal mammary development”. Breast Cancer Res. Treat. 47 (3): 201—8. PMID 9516076. S2CID 30440069. doi:10.1023/a:1005998832636.
- ^ a b v Kleinberg DL (1997). „Early mammary development: growth hormone and IGF-1”. J Mammary Gland Biol Neoplasia. 2 (1): 49—57. PMID 10887519. S2CID 41667675. doi:10.1023/A:1026373513521.
- ^ a b v Ruan W, Kleinberg DL (1999). „Insulin-like growth factor I is essential for terminal end bud formation and ductal morphogenesis during mammary development”. Endocrinology. 140 (11): 5075—81. PMID 10537134. doi:10.1210/endo.140.11.7095 .
- ^ a b v g d đ Kleinberg DL, Feldman M, Ruan W (2000). „IGF-I: an essential factor in terminal end bud formation and ductal morphogenesis”. J Mammary Gland Biol Neoplasia. 5 (1): 7—17. PMID 10791764. S2CID 25656770. doi:10.1023/A:1009507030633.
- ^ Kleinberg DL, Ruan W (2008). „IGF-I, GH, and sex steroid effects in normal mammary gland development”. J Mammary Gland Biol Neoplasia. 13 (4): 353—60. PMID 19034633. S2CID 24786346. doi:10.1007/s10911-008-9103-7.
- ^ Serra R, Crowley MR (2005). „Mouse models of transforming growth factor beta impact in breast development and cancer”. Endocr. Relat. Cancer. 12 (4): 749—60. PMID 16322320. doi:10.1677/erc.1.00936 .
- ^ LaMarca HL, Rosen JM (2007). „Estrogen regulation of mammary gland development and breast cancer: amphiregulin takes center stage”. Breast Cancer Res. 9 (4): 304. PMC 2206713 . PMID 17659070. doi:10.1186/bcr1740.
- ^ El-Attar HA, Sheta MI (2011). „Hepatocyte growth factor profile with breast cancer”. Indian J Pathol Microbiol. 54 (3): 509—13. PMID 21934211. doi:10.4103/0377-4929.85083 .
- ^ Coad, Jane; Dunstall, Melvyn (2011). Anatomy and Physiology for Midwives. Elsevier Health Sciences. str. 413—. ISBN 978-0-7020-3489-3.
- ^ Hynes, N. E.; Watson, C. J. (2010). „Mammary Gland Growth Factors: Roles in Normal Development and in Cancer”. Cold Spring Harbor Perspectives in Biology. 2 (8): a003186. ISSN 1943-0264. PMC 2908768 . PMID 20554705. doi:10.1101/cshperspect.a003186.
- ^ Harris, Jay R.; Lippman, Marc E.; C. Kent Osborne; Morrow, Monica (28. 3. 2012). Diseases of the Breast. Lippincott Williams & Wilkins. str. 94—. ISBN 978-1-4511-4870-1.
- ^ a b v Chong YM, Subramanian A, Sharma AK, Mokbel K (2007). „The potential clinical applications of insulin-like growth factor-1 ligand in human breast cancer”. Anticancer Res. 27 (3B): 1617—24. PMID 17595785.
- ^ Leonard R. Johnson (2003). Essential Medical Physiology. Academic Press. str. 770—. ISBN 978-0-12-387584-6.
- ^ a b Jernström H, Olsson H (1997). „Breast size in relation to endogenous hormone levels, body constitution, and oral contraceptive use in healthy nulligravid women aged 19–25 years”. Am. J. Epidemiol. 145 (7): 571—80. PMID 9098173. doi:10.1093/oxfordjournals.aje.a009153 .
- ^ Zhou J, Ng S, Adesanya-Famuiya O, Anderson K, Bondy CA (2000). „Testosterone inhibits estrogen-induced mammary epithelial proliferation and suppresses estrogen receptor expression”. FASEB J. 14 (12): 1725—30. PMID 10973921. S2CID 17172449. doi:10.1096/fj.99-0863com.
- ^ Lemaine V, Cayci C, Simmons PS, Petty P (2013). „Gynecomastia in adolescent males”. Semin Plast Surg. 27 (1): 56—61. PMC 3706045 . PMID 24872741. doi:10.1055/s-0033-1347166.
- ^ Sekhri, KK; Pitelka, DR; Deome, KB (septembar 1967). „Studies of mouse mammary glands. I. Cytomorphology of the normal mammary gland”. J Natl Cancer Inst. 39 (3): 459—90. PMID 6053715.
- ^ Hens, JR; Wysolmerski, J. J. (10. 8. 2005). „Key stages of mammary gland development: molecular mechanisms involved in the formation of the embryonic mammary gland”. Breast Cancer Res. 7 (5): 220—4. PMC 1242158 . PMID 16168142. doi:10.1186/bcr1306.
- ^ Makarem, M; Eaves, C. (april 2013). „Stem Cells and the Developing Mammary Gland”. J Mammary Gland Biol Neoplasia. 18 (2): 209—19. PMC 4161372 . PMID 23624881. doi:10.1007/s10911-013-9284-6.
- ^ Daniel, CW; Smith, GH (januar 1999). „The mammary gland: a model for development”. Journal of Mammary Gland Biology and Neoplasia. 4 (1): 3—8. PMID 10219902. S2CID 36670489. doi:10.1023/A:1018796301609.
- ^ Gudjonsson, T.; Rønnov-Jessen, L.; Villadsen, R.; Rank, F.; Bissell, M. J.; Petersen, O. W. (2002). „Normal and tumor-derived myoepithelial cells differ in their ability to interact with luminal breast epithelial cells for polarity and basement membrane deposition”. Journal of Cell Science. 115 (Pt 1): 39—50. PMC 2933194 . PMID 11801722. doi:10.1242/jcs.115.1.39.
- ^ Provenzano, P. P.; Inman, D. R.; Eliceiri, K. W.; Knittel, J. G.; Yan, L.; Rueden, C. T.; White, J. G.; Keely, P. J. (2008). „Collagen density promotes mammary tumor initiation and progression”. BMC Medicine. 6: 11. PMC 2386807 . PMID 18442412. doi:10.1186/1741-7015-6-11.

- ^ a b „With the Wild Things – Transcripts”. Digitalcollections.fiu.edu. Arhivirano iz originala 23. 3. 2013. g. Pristupljeno 05. 04. 2013.
- ^ a b Stockard, Mary (2005). „Raising Orphaned Baby Opossums”. Arhivirano iz originala 01. 07. 2010. g.. Alabama Wildlife Center.
- ^ Cunningham, Merle; LaTour, Mickey A.; Acker, Duane (2005). Animal Science and Industry. Pearson Prentice Hall. ISBN 978-0-13-046256-5.
- ^ Dog breeds vary in the number of mammary glands: larger breeds tend to have 5 pairs, smaller breeds have 4 pairs.[traži se izvor]
- ^ P Smith 2008 Red-Sided Short-Tailed Opossum. Fauna Paraguay
- ^ Julie Ann Mayer; Foley, John; Damon De La Cruz; Cheng-Ming Chuong; Widelitz, Randall (novembar 2008). „Conversion of the Nipple to Hair-Bearing Epithelia by Lowering Bone Morphogenetic Protein Pathway Activity at the Dermal-Epidermal Interface”. Am J Pathol. 173 (5): 1339—48. PMC 2570124 . PMID 18832580. doi:10.2353/ajpath.2008.070920.
- ^ Armati, Patricia J.; Dickman, Chris R.; Hume, Ian D. (17. 8. 2006). Marsupials. Cambridge University Press. ISBN 978-1-139-45742-2.
- ^ Kunz, T; Hosken, D (2009). „Male lactation: why, why not and is it care?”. Trends in Ecology & Evolution. 24 (2): 80—85. PMID 19100649. doi:10.1016/j.tree.2008.09.009.
- ^ Li, Peng; Knabe, Darrell A.; Kim, Sung Woo; Lynch, Christopher J.; Hutson, Susan M.; Wu, Guoyao (2009-08-01). „Lactating Porcine Mammary Tissue Catabolizes Branched-Chain Amino Acids for Glutamine and Aspartate Synthesis”. The Journal of Nutrition (na jeziku: engleski). 139 (8): 1502—1509. ISSN 0022-3166. PMC 3151199 . PMID 19549750. doi:10.3945/jn.109.105957 .
- ^ Oftedal, O. T. (2002). „The origin of lactation as a water source for parchment-shelled eggs”. Journal of Mammary Gland Biology and Neoplasia. 7 (3): 253—266. PMID 12751890. S2CID 8319185. doi:10.1023/A:1022848632125.
- ^ „Lactating on Eggs”. Arhivirano iz originala 19. 04. 2009. g.. Smithsonian National Zoo, 14 July 2003.
- ^ Oftedal, OT (2002). „The mammary gland and its origin during synapsid evolution”. Journal of Mammary Gland Biology and Neoplasia. 7 (3): 225—52. PMID 12751889. S2CID 25806501. doi:10.1023/A:1022896515287.
- ^ „Breast beginnings”. scienceblogs.com. Arhivirano iz originala 12. 03. 2007. g.
- ^ Vorbach, C.; Capecchi, M. R.; Penninger, J. M. (2006). „Evolution of the mammary gland from the innate immune system?”. BioEssays. 28 (6): 606—616. PMID 16700061. doi:10.1002/bies.20423.
- ^ Lefèvre, C. M.; Sharp, J. A.; Nicholas, K. R. (2010). „Evolution of Lactation: Ancient Origin and Extreme Adaptations of the Lactation System”. Annual Review of Genomics and Human Genetics. 11: 219—238. PMID 20565255. doi:10.1146/annurev-genom-082509-141806.
- ^ Graves, B. M.; Duvall, D. (1983). „A Role for Aggregation Pheromones in the Evolution of Mammallike Reptile Lactation”. The American Naturalist. 122 (6): 835. S2CID 84089647. doi:10.1086/284177.
Spoljašnje veze
uredi- „Anatomija mlečne žlezde”. Arhivirano iz originala 01. 12. 2005. g. od strane W. L. Hurley
- Anatomija grudi od strane Sir Astley Paston Cooper (1840) sa brojnim crtežima koji su u javnom vlasništvu.