Млечна жлезда
Млечна жлезда је ендокрина жлезда код људи и других сисара која производи млеко за исхрану младог потомства. Сисари су своје име добили по латинској речи mamma, „дојка“. Млечне жлезде су распоређене у органе као што су дојке код примата (на пример, људи и шимпанзе), виме код преживара (на пример краве, козе и јелени) и код још других животиња (на пример, паса и мачке). Галактореја, повремена производња млека у жлездама, може се јавити код било ког сисара, али код већине сисара, лактација, производња довољно млијека за његу, јавља се само код фенотипских женки које су гестовале последњих месеци или година. Усмеравају га хормоналне смјернице полних стероида. Код неколико врста сисара може доћи до мушке лактације. Код људи се мушка лактација може догодити само под одређеним околностима.
| Млечна жлезда | |
|---|---|
 Пресек млечне жлезде човека.
| |
| Детаљи | |
| Прекурзор | Мезодерм (крвних судова и везивног ткива) Ектодерм[3] (ћелијски елементи) |
| Артерија | Унутрашња торакална артерија Спољашња торакална артерија[1] |
| Вена | Унутрашља торакална вена Аксиларна вена[1] |
| Нерв | Супраклавикуларни нерви Интеркостални нерви[2] (бочне и медијалне гране) |
| Лимфа | Пекторални аксиларни лимфни чворови[1] |
| Идентификатори | |
| TA | A16.0.02.006 |
| FMA | 60088 |
| Анатомска терминологија | |
Сисари су подјељени у 3 групе: прототеријанци, метатеријанци и еутерије. У случају прототераца, и мушкарци и жене имају функционалне млечне жлезде, али њихове млечне жлезде су без брадавица. Ове млечне жлезде су модификоване лојне жлезде. Што се тиче метатеријанаца и еутерија, само жене имају функционалне млечне жлезде. Њихове млечне жлезде могу се назвати дојкама или вименом. У случају дојки, свака млечна жлезда има своју брадавицу. У случају вимена, парови млечних жлезда чине једну масу, а на њу виси више од једне брадавице. На пример, краве и биволи имају по једно виме са четири сисе, док овце и козе имају по две сисе који вире из вимена. Ове млечне жлезде су модификоване знојне жлезде.
Структура
уредиОсновне компоненте зреле млечне жлезде су алвеоле (велике неколико милиметара), које су обложене кубоидним ћелијама које луче млеко и окружене миоепителним ћелијама. Ове алвеоле се спајају у групе познате као лобули. Свака лобула има млечни канал који се одводи у отворе на брадавици. Миоепителне ћелије се контрахују под стимулацијом окситоцина, излучујући млеко које луче алвеоларне јединице у лумен лобула према брадавици. Како дојенче почиње сисати, долази до „рефлекса пуштања“ посредованог окситоцином, а мајчино млеко се излучује - а не усисава - из жлезде у бебина уста. [4]
Сво ткиво које лучи млеко које води до једног млечног канала назива се заједнички „једноставна млечна жлезда“; у „сложеној млечној жлезди“ све једноставне млечне жлезде служе једној брадавици. Људи обично имају две сложене млечне жлезде, по једну у свакој дојци, а свака сложена млечна жлезда састоји се од 10–20 једноставних жлезда. Присуство више од две брадавице познато је као полителија, а присуство више од две сложене млечне жлезде као полимастија.
Одржавање тачне поларизоване морфологије стабла млечног канала захтева још једну битну компоненту - екстрацелуларни матрикс млечних епителних ћелија (ЕЦМ), који заједно са адипоцитима, фибробластом, инфламаторним ћелијама и другим чине млечну строму. [5] ЕЦМ млечног епитела углавном садржи миоепителну базалну мембрану и везивно ткиво. Они не само да помажу у подршци основне структуре млечних жлезда, већ и служе као комуникациони мост између млечних епитела и њиховог локалног и глобалног окружења током целог развоја овог органа. [6][7]
Хистологија
уреди
Млечна жлезда је специфична врста апокрине жлезде специјализована за производњу колострума током порођаја. Многи извори тврде да су млечне жлезде модификоване знојне жлезде. [8][9][10] Неки аутори то оспоравају и уместо тога тврде да су лојне жлезде. [8]
Развој
уредиМлечне жлезде се развијају током различитих циклуса раста. Постоје у оба пола током ембрионалне фазе, формирајући само рудиментарно дрво канала при рођењу. У овој фази развој млечних жлезда зависи од системских (и мајчиних) хормона, [5] али је такође под (локалном) регулацијом паракрине комуникације између суседних епителних и мезенхималних ћелија протеинима повезаним са паратироидним хормонима PTHrP. [11] Овај локално секретовани фактор доводи до низа позитивних повратних информација изнутра између ове две врсте ћелија, тако да епителне ћелије млечних пупољака могу да се размножавају и ничу у мезенхимални слој док не дођу до масног слоја да би започеле први круг гранања. [5] Истовремено, ембрионалне мезенхимске ћелије око епителног пупољка добијају факторе секреције које активира PTHrP, као што је BMP4. Ове мезенхимске ћелије могу се трансформисати у густи, специфични за млечне жлезде мезенхим, који се касније развија у везивно ткиво са влакнастим нитима, формирајући крвне судове и лимфни систем. [12] Базална мембрана, која углавном садржи ламинин и колаген, настала након тога диференцираним миоепителним ћелијама, задржава поларитет овог примарног стабла канала. Ове компоненте екстрацелуларног матрикса снажне су одреднице морфогенезе канала. [13]
Биохемија
уредиЕстроген и хормон раста (ГХ) су од суштинске важности за дукталну компоненту развоја млечних жлезда и делују синергијски да би то посредовали. [14][15][16][17][18] Ни естроген ни ГХ нису способни да индукују дуктални развој један без другог.[15][16][17][18][15][16][17][18][19] Утврђено је да је улога ГХ у дукталном развоју углавном посредована његовом индукцијом секреције инсулину сличног фактора раста (ИГФ-1), који се јавља системски (углавном пореклом из јетре). [18] Међутим, сам ГХ такође делује независно од ИГФ-1 да стимулише развој дуктала регулацијом експресије естрогенског рецептора (ЕР) у ткиву млечне жлезде, што је низводни ефекат активације ГХР млечне жлезде. [18] У сваком случају, за разлику од ИГФ-1, сам ГХ није битан за развој млечних жлезда, а ИГФ-1 у спрези са естрогеном може да индукује нормалан развој млечних жлезда без присуства ГХ. [18] Поред ИГФ-1, присутни су и други паракрини фактори раста као што су епидермални фактор раста (ЕГФ), трансформишући фактор раста бета (ТГФ-β), [20] амфирегулин, [21] фактор раста фибробласта (ФГФ) и фактор раста хепатоцита ( ХГФ) [22] су укључени у развој дојке као посредници низводно од полних хормона и ГХ / ИГФ-1. [23][24][25]
Током ембрионалног развоја, ниво ИГФ-1 је низак и постепено расте од рођења до пубертета. [26] У пубертету нивои ГХ и ИГФ-1 достижу највиши ниво у животу и естроген почиње да се лучи у великим количинама код жена, тада се углавном одвија дуктални развој. [26] Под утицајем естрогена расте и стромално и масно ткиво које окружује дуктални систем млечних жлезда. [27] После пубертета, нивои ГХ и ИГФ-1 се прогресивно смањују, што ограничава даљи развој до трудноће, ако се догоди.[тражи се извор] Током трудноће, прогестерон и пролактин су од суштинске важности за посредовање лобулоалвеоларног развоја у ткиву млечне жлезде препуној естрогену, што се дешава у припреми лактације и неге. [26].[14][28]
Андрогени као што је тестостерон инхибирају развој млечних жлезда посредованих естрогеном (нпр. Смањењем локалне експресије ЕР) активирањем андрогених рецептора изражених у ткиву млечних жлезда, [28][29] и заједно са релативно ниским нивоима естрогена, узрок су недостатак развијених млечних жлезда код мушкараца. [30]
Преглед
уредиПре рођења
уредиРазвој млечних жлезда карактерише јединствени процес којим епител напада инвазију строме. Развој млечне жлезде јавља се углавном након рођења. Током пубертета, формирање тубула је повезано са разгранатом морфогенезом која успоставља основну арбореалну мрежу канала који излазе из брадавице. [31]
У развоју, епител млечне жлезде непрестано производе и одржавају ретке епителне ћелије, назване родовима млечних жлезда, за које се на крају сматра да потичу од матичних ћелија настањених у ткиву.
Развој ембрионалне млечне жлезде може се поделити у низ специфичних фаза. У почетку се формирање млечних линија које се протежу између предњих и задњих удова обострано на свакој страни средње линије дешава око ембрионалног дана 10.5 (Е10.5). Друга фаза се дешава на Е11.5 када започиње формирање плакова дуж млечне линије млека. Ово ће на крају довести до појаве брадавице. На крају, трећа фаза се дешава на Е12.5 и укључује инвагинацију ћелија унутар плакова у мезенхим, што доводи до анлаже млечних жлезда. [32]
Примитивне (матичне) ћелије се откривају у ембриону и њихов број се непрестано повећава током развоја [33]
Раст
уредиПостнатално, млечни канали се издужују у млечни јастучић - млечне жлезде. Затим, почев од око четири недеље старости, раст дукталног млечног млека значајно се повећава са продирањем канала према лимфном чвору. Крајњи пупољци терминала, високо пролиферативне структуре које се налазе на врховима упадних канала, током ове фазе се увелико шире и повећавају. Овај развојни период карактерише појава крајњих пупољака и траје до старости око 7-8 недеља.
У пубертетском стадијуму млечни канали су инвадирали до краја млечне масне подлоге. У овом тренутку крајњи пупољци постају мање пролиферативни и смањују се у величини. Бочне гране се формирају из примарних канала и почињу да испуњавају млечни јастучић млечне жлезде. Дуктални развој опада доласком полне зрелости и пролази кроз естроусне циклусе (проеструс, еструс, метеструс и диеструс). Као резултат естроусног циклуса, млечна жлезда се подвргава динамичким променама где ћелије пролиферирају, а затим регресирају на уређен начин. [34]
Трудноћа
уредиТоком трудноће, дуктални системи се брзо шире и формирају алвеоларне структуре унутар грана које ће се користити за производњу млека. Након порођаја, лактација се јавља унутар млечне жлезде; лактација укључује лучење млека луминалним ћелијама у алвеолама. Контракција миоепителних ћелија које окружују алвеоле довешће до избацивања млека кроз канале и у брадавицу за дојенче. Након одбијања дојенчета, лактација престаје и млечна жлезда се окреће сама себи, процес који се назива инволуција. Овај процес укључује контролисани колапс епителних ћелија млечне жлезде где ћелије контролисано започињу апоптозу, враћајући млечну жлезду назад у пубертетско стање.
Постменопауза
уредиТоком постменопаузе, због много нижих нивоа естрогена и због нижих нивоа ГХ и ИГФ-1, који се смањују са годинама, ткиво млечних жлезда атрофира и млечне жлезде постају све мање.
Клинички значај
уредиТуморигенеза млечних жлезда може бити индикована биохемијски неправилним нивоом експресије хормона у циркулацији или локалних компонената ЕЦМ, [35] или механичком променом напетости млечне строме. [36] У било којој од ове две околности, епителне ћелије млека ће расти ван контроле и на крају резултирати раком. Скоро сви случајеви рака дојке потичу из лобула или канала млечних жлезда.
Остали сисари
уредиГенерално
уредиГруди одрасле жене разликују се од већине осталих сисара који имају мање видљиве млечне жлезде. Број и положај млечних жлезда варира код различитих сисара. Истурене груди и пратеће жлезде могу се налазити било где дуж две линије млека. Генерално, већина сисара развија млечне жлезде у паровима дуж ових линија, с бројем који приближно одговара броју младих типично рођених у исто време. Број сиса варира од 2 (код већине примата) до 18 (код свиња). Амерички опосум има 13, један од ретких сисара са непарним бројем.[37][38]Следећа табела наводи број и положај сисара и жлезда нађених код сисара:
| Врсте[39] | Предњи (торакално) |
Средњи (абноминално) |
Иза (ингвинално) |
Укупно |
|---|---|---|---|---|
| Коза, Овца, Коњ Морско прасе |
0 | 0 | 2 | 2 |
| Говедо | 0 | 0 | 4 | 4 |
| Мачка | 2 | 2 | 4 | 8 |
| Пас[40] | 4 | 2 | 2 или 4 | 8 или 10 |
| Миш | 6 | 0 | 4 | 10 |
| Пацов | 6 | 2 | 4 | 12 |
| Свиња | 6 | 6 | 6 | 18 |
| Сурлаши, примати | 2 | 0 | 0 | 2 |
| Амерички опосум[37][38] | 0 | 0 | 13 | 13 |
| Јужни црвенобоки опосум[41] | 0 | 0 | 25 или 27 | 25 или 27 |
Мушки сисари обично имају рудиментарне млечне жлезде и брадавице, уз неколико изузетака: мушки мишеви немају брадавице, [42] мушки торбари немају млечне жлезде, [43] а мушки коњи немају брадавице и млечне жлезде. Мушка лактација се ретко јавља код неких врста. [44]
Млечне жлезде су праве фабрике протеина, [45] а неколико лабораторија је изградило трансгене животиње, углавном козе и краве, како би производиле протеине за фармацеутску употребу. Комплексни гликопротеини, као што су моноклонска антитела или антитромбин, не могу да произведу генетски модификоване бактерије, а производња код живих сисара је много јефтинија од употребе ћелијских култура сисара.
Еволуција
уредиПостоји много теорија о томе како су млечне жлезде еволуирале. На пример, сматра се да је млечна жлезда трансформисана знојна жлезда, ближе повезана са апокриним знојним жлездама. [46] Будући да млечне жлезде не фосилизују добро, тешко је подржати такве теорије фосилним доказима. Многе тренутне теорије заснивају се на поређењу између линија живих сисара - монотрема, торбарских животиња и еутерија. Једна теорија предлаже да су млечне жлезде еволуирале из жлезда које су коришћене за одржавање влажних јајашаца раних сисара [47][48] и без заразе [49][50] (монотреми још увек полажу јаја). Друге теорије сугеришу да су рани секрет директно користили излежени млади [51] или да су га млади користили да би им помогао да се оријентишу према мајкама. [52]
Сматра се да се лактација развила много пре еволуције млечне жлезде и сисара.
Додатне слике
уреди-
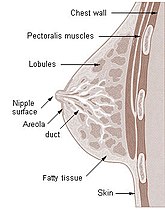 Пресек дојке жене
Пресек дојке жене -

-

-

-

-

-








Види још
уредиРеференце
уреди- ^ а б в Macéa, José Rafael; Fregnani, José Humberto Tavares Guerreiro (1. 12. 2006). „Anatomy of the Thoracic Wall, Axilla and Breast” (PDF). International Journal of Morphology. 24 (4). doi:10.4067/S0717-95022006000500030
 .
.
- ^ Lawrence, Ruth A.; Lawrence, Robert M. (2010-09-30). Breastfeeding: A Guide for the Medical Profession (7th изд.). Maryland Heights, Maryland: Mosby/Elsevier. стр. 54. ISBN 9781437735901.
- ^ Gray, Henry (1918). Anatomy of the Human Body.
- ^ Newton, Michael; Newton, Niles Rumely (децембар 1948). „The let-down reflex in human lactation”. The Journal of Pediatrics. 33 (6): 698—704. doi:10.1016/S0022-3476(48)80075-2.
- ^ а б в Watson, C. J.; Khaled, W. T. (2008). „Mammary development in the embryo and adult: A journey of morphogenesis and commitment”. Development. 135 (6): 995—1003. PMID 18296651. doi:10.1242/dev.005439 .
- ^ Wiseman, B. S.; Werb, Z. (2002). „Stromal Effects on Mammary Gland Development and Breast Cancer”. Science. 296 (5570): 1046—1049. Bibcode:2002Sci...296.1046W. PMC 2788989 . PMID 12004111. doi:10.1126/science.1067431.
- ^ Pavlovich, A. L.; Manivannan, S.; Nelson, C. M. (2010). „Adipose Stroma Induces Branching Morphogenesis of Engineered Epithelial Tubules”. Tissue Engineering Part A. 16 (12): 3719—3726. PMC 2991209 . PMID 20649458. doi:10.1089/ten.TEA.2009.0836.
- ^ а б Ackerman (2005) ch.1 „Apocrine Units”. Архивирано из оригинала 21. 04. 2011. г.
- ^ Moore (2010) ch.1 Thorax, p. 99
- ^ Krstic, Radivoj V. (18. 3. 2004). Human Microscopic Anatomy: An Atlas for Students of Medicine and Biology. Springer. стр. 466. ISBN 9783540536666.
- ^ Wysolmerski, J. J.; Philbrick, W. M.; Dunbar, M. E.; Lanske, B.; Kronenberg, H.; Broadus, A. E. (1998). „Rescue of the parathyroid hormone-related protein knockout mouse demonstrates that parathyroid hormone-related protein is essential for mammary gland development”. Development. 125 (7): 1285—1294. PMID 9477327. doi:10.1242/dev.125.7.1285.
- ^ Hens, J. R.; Wysolmerski, J. J. (2005). „Key stages of mammary gland development: Molecular mechanisms involved in the formation of the embryonic mammary gland”. Breast Cancer Research. 7 (5): 220—224. PMC 1242158 . PMID 16168142. doi:10.1186/bcr1306.
- ^ Montévil, Maël; Speroni, Lucia; Sonnenschein, Carlos; Soto, Ana M. (2016-10-01). „Modeling mammary organogenesis from biological first principles: Cells and their physical constraints”. Progress in Biophysics and Molecular Biology. From the Century of the Genome to the Century of the Organism: New Theoretical Approaches. 122 (1): 58—69. PMC 5563449 . PMID 27544910. doi:10.1016/j.pbiomolbio.2016.08.004.
- ^ а б Brisken; Malley (2. 12. 2010). „Hormone Action in the Mammary Gland”. Cold Spring Harbor Perspectives in Biology. 2 (12): a003178. PMC 2982168 . PMID 20739412. doi:10.1101/cshperspect.a003178.
- ^ а б в Kleinberg DL (1998). „Role of IGF-I in normal mammary development”. Breast Cancer Res. Treat. 47 (3): 201—8. PMID 9516076. S2CID 30440069. doi:10.1023/a:1005998832636.
- ^ а б в Kleinberg DL (1997). „Early mammary development: growth hormone and IGF-1”. J Mammary Gland Biol Neoplasia. 2 (1): 49—57. PMID 10887519. S2CID 41667675. doi:10.1023/A:1026373513521.
- ^ а б в Ruan W, Kleinberg DL (1999). „Insulin-like growth factor I is essential for terminal end bud formation and ductal morphogenesis during mammary development”. Endocrinology. 140 (11): 5075—81. PMID 10537134. doi:10.1210/endo.140.11.7095 .
- ^ а б в г д ђ Kleinberg DL, Feldman M, Ruan W (2000). „IGF-I: an essential factor in terminal end bud formation and ductal morphogenesis”. J Mammary Gland Biol Neoplasia. 5 (1): 7—17. PMID 10791764. S2CID 25656770. doi:10.1023/A:1009507030633.
- ^ Kleinberg DL, Ruan W (2008). „IGF-I, GH, and sex steroid effects in normal mammary gland development”. J Mammary Gland Biol Neoplasia. 13 (4): 353—60. PMID 19034633. S2CID 24786346. doi:10.1007/s10911-008-9103-7.
- ^ Serra R, Crowley MR (2005). „Mouse models of transforming growth factor beta impact in breast development and cancer”. Endocr. Relat. Cancer. 12 (4): 749—60. PMID 16322320. doi:10.1677/erc.1.00936 .
- ^ LaMarca HL, Rosen JM (2007). „Estrogen regulation of mammary gland development and breast cancer: amphiregulin takes center stage”. Breast Cancer Res. 9 (4): 304. PMC 2206713 . PMID 17659070. doi:10.1186/bcr1740.
- ^ El-Attar HA, Sheta MI (2011). „Hepatocyte growth factor profile with breast cancer”. Indian J Pathol Microbiol. 54 (3): 509—13. PMID 21934211. doi:10.4103/0377-4929.85083 .
- ^ Coad, Jane; Dunstall, Melvyn (2011). Anatomy and Physiology for Midwives. Elsevier Health Sciences. стр. 413—. ISBN 978-0-7020-3489-3.
- ^ Hynes, N. E.; Watson, C. J. (2010). „Mammary Gland Growth Factors: Roles in Normal Development and in Cancer”. Cold Spring Harbor Perspectives in Biology. 2 (8): a003186. ISSN 1943-0264. PMC 2908768 . PMID 20554705. doi:10.1101/cshperspect.a003186.
- ^ Harris, Jay R.; Lippman, Marc E.; C. Kent Osborne; Morrow, Monica (28. 3. 2012). Diseases of the Breast. Lippincott Williams & Wilkins. стр. 94—. ISBN 978-1-4511-4870-1.
- ^ а б в Chong YM, Subramanian A, Sharma AK, Mokbel K (2007). „The potential clinical applications of insulin-like growth factor-1 ligand in human breast cancer”. Anticancer Res. 27 (3B): 1617—24. PMID 17595785.
- ^ Leonard R. Johnson (2003). Essential Medical Physiology. Academic Press. стр. 770—. ISBN 978-0-12-387584-6.
- ^ а б Jernström H, Olsson H (1997). „Breast size in relation to endogenous hormone levels, body constitution, and oral contraceptive use in healthy nulligravid women aged 19–25 years”. Am. J. Epidemiol. 145 (7): 571—80. PMID 9098173. doi:10.1093/oxfordjournals.aje.a009153 .
- ^ Zhou J, Ng S, Adesanya-Famuiya O, Anderson K, Bondy CA (2000). „Testosterone inhibits estrogen-induced mammary epithelial proliferation and suppresses estrogen receptor expression”. FASEB J. 14 (12): 1725—30. PMID 10973921. S2CID 17172449. doi:10.1096/fj.99-0863com.
- ^ Lemaine V, Cayci C, Simmons PS, Petty P (2013). „Gynecomastia in adolescent males”. Semin Plast Surg. 27 (1): 56—61. PMC 3706045 . PMID 24872741. doi:10.1055/s-0033-1347166.
- ^ Sekhri, KK; Pitelka, DR; Deome, KB (септембар 1967). „Studies of mouse mammary glands. I. Cytomorphology of the normal mammary gland”. J Natl Cancer Inst. 39 (3): 459—90. PMID 6053715.
- ^ Hens, JR; Wysolmerski, J. J. (10. 8. 2005). „Key stages of mammary gland development: molecular mechanisms involved in the formation of the embryonic mammary gland”. Breast Cancer Res. 7 (5): 220—4. PMC 1242158 . PMID 16168142. doi:10.1186/bcr1306.
- ^ Makarem, M; Eaves, C. (април 2013). „Stem Cells and the Developing Mammary Gland”. J Mammary Gland Biol Neoplasia. 18 (2): 209—19. PMC 4161372 . PMID 23624881. doi:10.1007/s10911-013-9284-6.
- ^ Daniel, CW; Smith, GH (јануар 1999). „The mammary gland: a model for development”. Journal of Mammary Gland Biology and Neoplasia. 4 (1): 3—8. PMID 10219902. S2CID 36670489. doi:10.1023/A:1018796301609.
- ^ Gudjonsson, T.; Rønnov-Jessen, L.; Villadsen, R.; Rank, F.; Bissell, M. J.; Petersen, O. W. (2002). „Normal and tumor-derived myoepithelial cells differ in their ability to interact with luminal breast epithelial cells for polarity and basement membrane deposition”. Journal of Cell Science. 115 (Pt 1): 39—50. PMC 2933194 . PMID 11801722. doi:10.1242/jcs.115.1.39.
- ^ Provenzano, P. P.; Inman, D. R.; Eliceiri, K. W.; Knittel, J. G.; Yan, L.; Rueden, C. T.; White, J. G.; Keely, P. J. (2008). „Collagen density promotes mammary tumor initiation and progression”. BMC Medicine. 6: 11. PMC 2386807 . PMID 18442412. doi:10.1186/1741-7015-6-11.

- ^ а б „With the Wild Things – Transcripts”. Digitalcollections.fiu.edu. Архивирано из оригинала 23. 3. 2013. г. Приступљено 05. 04. 2013.
- ^ а б Stockard, Mary (2005). „Raising Orphaned Baby Opossums”. Архивирано из оригинала 01. 07. 2010. г.. Alabama Wildlife Center.
- ^ Cunningham, Merle; LaTour, Mickey A.; Acker, Duane (2005). Animal Science and Industry. Pearson Prentice Hall. ISBN 978-0-13-046256-5.
- ^ Dog breeds vary in the number of mammary glands: larger breeds tend to have 5 pairs, smaller breeds have 4 pairs.[тражи се извор]
- ^ P Smith 2008 Red-Sided Short-Tailed Opossum. Fauna Paraguay
- ^ Julie Ann Mayer; Foley, John; Damon De La Cruz; Cheng-Ming Chuong; Widelitz, Randall (новембар 2008). „Conversion of the Nipple to Hair-Bearing Epithelia by Lowering Bone Morphogenetic Protein Pathway Activity at the Dermal-Epidermal Interface”. Am J Pathol. 173 (5): 1339—48. PMC 2570124 . PMID 18832580. doi:10.2353/ajpath.2008.070920.
- ^ Armati, Patricia J.; Dickman, Chris R.; Hume, Ian D. (17. 8. 2006). Marsupials. Cambridge University Press. ISBN 978-1-139-45742-2.
- ^ Kunz, T; Hosken, D (2009). „Male lactation: why, why not and is it care?”. Trends in Ecology & Evolution. 24 (2): 80—85. PMID 19100649. doi:10.1016/j.tree.2008.09.009.
- ^ Li, Peng; Knabe, Darrell A.; Kim, Sung Woo; Lynch, Christopher J.; Hutson, Susan M.; Wu, Guoyao (2009-08-01). „Lactating Porcine Mammary Tissue Catabolizes Branched-Chain Amino Acids for Glutamine and Aspartate Synthesis”. The Journal of Nutrition (на језику: енглески). 139 (8): 1502—1509. ISSN 0022-3166. PMC 3151199 . PMID 19549750. doi:10.3945/jn.109.105957 .
- ^ Oftedal, O. T. (2002). „The origin of lactation as a water source for parchment-shelled eggs”. Journal of Mammary Gland Biology and Neoplasia. 7 (3): 253—266. PMID 12751890. S2CID 8319185. doi:10.1023/A:1022848632125.
- ^ „Lactating on Eggs”. Архивирано из оригинала 19. 04. 2009. г.. Smithsonian National Zoo, 14 July 2003.
- ^ Oftedal, OT (2002). „The mammary gland and its origin during synapsid evolution”. Journal of Mammary Gland Biology and Neoplasia. 7 (3): 225—52. PMID 12751889. S2CID 25806501. doi:10.1023/A:1022896515287.
- ^ „Breast beginnings”. scienceblogs.com. Архивирано из оригинала 12. 03. 2007. г.
- ^ Vorbach, C.; Capecchi, M. R.; Penninger, J. M. (2006). „Evolution of the mammary gland from the innate immune system?”. BioEssays. 28 (6): 606—616. PMID 16700061. doi:10.1002/bies.20423.
- ^ Lefèvre, C. M.; Sharp, J. A.; Nicholas, K. R. (2010). „Evolution of Lactation: Ancient Origin and Extreme Adaptations of the Lactation System”. Annual Review of Genomics and Human Genetics. 11: 219—238. PMID 20565255. doi:10.1146/annurev-genom-082509-141806.
- ^ Graves, B. M.; Duvall, D. (1983). „A Role for Aggregation Pheromones in the Evolution of Mammallike Reptile Lactation”. The American Naturalist. 122 (6): 835. S2CID 84089647. doi:10.1086/284177.
Спољашње везе
уреди- „Анатомија млечне жлезде”. Архивирано из оригинала 01. 12. 2005. г. од стране W. L. Hurley
- Анатомија груди од стране Sir Astley Paston Cooper (1840) са бројним цртежима који су у јавном власништву.