Спавање
Спавање је природно, периодично и физиолошки реверзибилно стање умањене будности, које карактеришу смањена перцепција и реаговање на спољашње дражи. Оно је праћено комплексним бихевиоралним (хркање, мрмљање, окретање у кревету итд.) и биолошким процесима (измена срчане фреквенције, дисања, лучења хормона итд.).[1] Може се дефинисати и као несвесно стање из кога се особа може пробудити сензорним или другим дражима, и у том смислу се разликује од коме, која је такође несвесно стање, али се из ње на овај начин не може пробудити.

Спавање је универзални облик понашања који се јавља код свих животињских врста, од инсеката до сисара.
Иако спавање већ вековима фасцинира људе и предмет је константног истраживања од стране многих научних дисциплина, његове физиолошке функције нису сасвим познате и у великој мери се могу подвести под хипотезе, иако човек спавајући проведе приближно трећину живота. Оно је од посебне важности за психијатрију јер се готово све психијатријске болести између осталог манифестују неким обликом поремећаја спавања, па су ови поремећаји често у склопу стандардизованих дијагностичких критеријума за одређене психијатријске болести.[2]
Неурофизиологија циркадијалног ритма и спавања
уреди
Биолошке функције спавања нису потпуно разјашњене. Зна се да оно настаје у мозгу и да постоји због мозга, јер је то једини орган за који је утврђено да има користи од спавања. Сматра се да је његова најбитнија улога ресторативна, што обухвата опоравак интегритета неурона и матурацију синапси, чиме се омогућава консолидација памћења и олакшава учење. Нервни центри за спавање нису јасно анатомски дефинисани, већ се састоје од неколико разбацаних група неурона. Неурони укључени у започињање и одржавање спороталасног спавања налазе се у ретикуларној формацији продужене мождине, таламусу, хипоталамусу и медијалним базалним једрима теленцефалона, док се неурони укључени у започињање и одржавање будности налазе у ретикуларној формацији мезенцефалона, субталамусу, таламусу и медијалним базалним једрима теленцефалона. Мала група холинергичких неурона који учествују у РЕМ спавању додатно је груписана у горњем делу ретикуларне формације можданог моста (понса) (врсте спавања ће детаљније бити описане у истоименом одељку).[1] Штавише, верује се како управо та група нервних ћелија, која се назива фронтално поље џиновских ћелија (ФТГ), директно иницира РЕМ спавање. Доказано је да стимулација ретикуларне формације средњег мозга, која се назива асцендентни ретикуларни активирајући систем (АРАС), изазива буђење животиње која спава и разарање овог система проузрокује коматозно стање, а сличан ефекат има и стимулација задњег хипоталамуса, док стимулација предњег хипоталамуса узрокује спавање. Заправо, будно стање зависи управо од АРАС-а, који шаље импулсе преко релејних једара таламуса до коре великог мозга, а управо прилив ових импулса доводи до физиолошких измена кортикалних активности, тј. кортикалног тонуса, који је основа свих когнитивних и моторних функција. Према класичној, тзв. пасивној теорији спавања, спавање је пасиван процес, односно стање општег смањења неуронске активности и смањеног прилива дражи, због чега се АРАС налази у стању пасивне инхибиције. Испоставило се, међутим, да се ради о много сложенијем процесу, па тако стимулација понтиних рафе једара, предњег дела хипоталамуса и орбиталног дела фронталног кортекса такође може изазвати НРЕМ спавање. Неурони асцендентног ретикуларног активацијског система луче ацетилхолин, норадреналин или серотонин; неурони предњег хипоталамуса гама-аминобутерну киселину (ГАБА), а неурони задњег хипоталамуса хистамин. Према важећој, активној теорији спавања, прелазак из стања спавања у стање будности и обрнуто узрокован је небалансираним реципроцитетом у активности неурона у АРАС, па када су доминантно активни норадреналински (у локусу церулеусу) и серотонински (у рафе једрима) неурони долази до смањене активације ацетилхолинских неурона (у ретикуларној формацији понса), што доводи до стања будности. Ако је баланс активације обрнут, тј. у корист ацетилхолинских неурона, долази до РЕМ спавања. Такође, појачано лучење ГАБА и смањено лучење хистамина узрокује НРЕМ спавање смањујући активност мождане коре и таламуса. Када се лучење ГАБА смањи, а хистаминско повећа, наступа будност.[3]

Регулаторни биолошки механизми у великој мери имају променљив, осцилаторни тип контроле важних временски зависних физиолошких функција. Живот на Земљи одвија се у складу са бројним ритмичким променама. Температура, количина падавина и трајање дана и ноћи мењају се са променом годишњих доба (кретање Земље око Сунца); свакодневно долази до смењивања циклуса дана и ноћи (кретање Земље око своје осе), као и плиме и осеке (кретање Месеца око Земље). Како би се успешно адаптирале и опстале, животиње морају прилагодити своје понашање цикличним променама средине у којој живе.[4] Ове цикличне промене активности организма настале услед утицаја периодичних промена спољних фактора називају се биолошки ритмови. Циклус спавања и будности једна је од функција људског организма које се смењују у складу са циркадијалном периодичношћу, односно везана је за спољне промене циклуса светла и мрака. Нормална периодичност циклуса износи нешто преко 24 сата, али се може пореметити ако се особа изолује од утицаја смењивања светлости и мрака или промени временску зону. Структуре одговорне за контролу ендогеног (унутрашњег) 24-часовног циркадијалног ритма су супрахијазматска једра хипоталамуса, која се налазе са обе стране изнад оптичке хијазме (раскрснице). Њихови неурони, иако нису део ретикуларног активационог система, такође играју битну улогу у контроли ритма спавања и будности. Ова једра примају информације о смењивању циклуса светла и мрака путем ретинохипоталамичких влакана директно из мрежњаче и као одговор узрокују нервне и хуморалне сигнале који усклађују бројне биолошке ритмове. Она делују на одговарајуће неуроне кичмене мождине, који даље утичу на активност неурона у горњем цервикалном ганглиону, чији се аксони пројектују у пинеалну жлезду (епифизу). Као одговор на нервне сигнале из горњег цервикалног ганглиона, епифиза лучи хормон мелатонин. Мелатонин се појачано синтетише и лучи при мрачном окружењу, а врло мало при светлом, па се сматра да он координише промене које настају због смењивања циклуса светла и мрака. Његово лучење је могуће само у условима природног мрака, а не када је, на пример, соба замрачена у току дана. Максимално лучење овог хормона достиже се између 2 и 4 сата ујутро.[1] Разарање супрахијазматских једара доводи до поремећаја циклуса спавања и будности.
Циркадијални ритам спавања човека састоји се од око 8 сати спавања и 16 сати будности.
Неуроендокрине промене током спавања
уредиПостоји комплексна веза између хормона и спавања. Неки хормони могу утицати на спавање, као што спавање може утицати на лучење појединих хормона. Поред мелатонина, смењивање циклуса спавања и будности утиче и на секрецију других хормона који делују у складу са циркадијалним променама. Хормон раста се највише лучи раније током спавања, а његову секрецију подстиче спороталасно спавање. Спавање такође утиче на појачано лучење пролактина, мада оно свој врхунац достиже касније него хормон раста, негде средином ноћи. Међутим, таласи хормона раста и пролактина могу се спорадично лучити и на самом почетку спавања, без обзира на време када се спава. Верује се да ова два хормона делују на процес спавања механизмом позитивне повратне спреге, па тако хормон раста стимулише спороталасно, док пролактин повећава РЕМ спавање. Са друге стране, лучење тиреостимулишућег хормона достиже врхунац увече, пре самог почетка спавања, док је током самог спавања инхибисано, а додатно га подстиче продужена будност. Хипоталамо-хипофизно-надбубрежна осовина је такође најнеактивнија на почетку ноћног спавања. На почетку спавања се инхибише и лучење кортизола, док се његов ниво, као и ниво адренокортикотропина повећавају при крају спавања, непосредно пре буђења, па је вероватно како ова два хормона учествују у изазивању будности. Лоша хигијена спавања и продужена будност могу имати утицај на ендокрини систем, па на пример, пацијенти са опструктивном апнејом у спавању луче мање количине хормона раста и пролактина, а при дужем неспавању увече долази до појачане активности хипоталамо-хипофизно-надбубрежне осовине, која је у нормалним условима тада мање активна.
Врсте спавања
уредиСпавање је комплексан процес, састављен од више фаза, од којих свака има специфичне карактеристике. Током њега долази до различитих биолошких и бихевиоралних промена и догађаја који су у вези са одговарајућим фазама спавања. Бихевиоралне догађаје, као што је мрмљање или окретање у кревету, могуће је непосредно посматрати, док се за регистровање физиолошких (првенствено неурофизиолошких) промена током спавања у научне или дијагностичке сврхе користи полисомнографија. То је неинвазивна метода којом се током (обично целовечерњег) спавања мере различити физиолошки параметри. Основни параметри који се прате у неурофизиолошким испитивањима (научноистраживачким и клиничким) јесу електроенцефалограм (ЕЕГ), електроокулограм (ЕОГ; мерење активности покрета очију) и површински електромиограм (ЕМГ; испитивање електричне мишићне активности) мишића у пределу браде и врата. Такође, у зависности од циља обављања полисомнографије, прате се и други параметри (ЕКГ, засићеност крви кисеоником, положај тела, интензитет дисања, покрети екстремитета...), који се могу комбиновати и са подацима о понашању особа током спавања добијеним аудио и видео технологијом, како би се дошло до што прецизнијих и обухватнијих информација.
Електроенцефалографске карактеристике будности и спавања
уреди
Електричне промене у неуронима мозга, посебно у оним лоцираним ближе његовој површини, могу се регистровати преко електрода спојених за површину лобање путем сланог гела који проводи електрицитет. Ове електроде региструју промене и шаљу их до електроенцефалографске машине. Запис који ова машина генерише зове се електроенцефалограм, који у ствари представља разлику у потенцијалу између две тачке на површини лобање.[1] Електричне промене региструју се у виду можданих таласа.
Најзаступљенији таласи у будне, опуштене особе са затвореним очима су алфа-таласи, фреквенције од 8 до 13 Хз и волтаже од око 50 микроволти. Ови таласи су повезани са смањеним нивоом пажње и када се они генеришу особа обично описује да се осећа опуштено и срећно, а најчешће се региструју са потиљачног (окципиталног), а затим са чеоног (фронталног) и теменог (паријеталног) режња.[5]
Када људи усмере пажњу на неки спољни објекат или интензивно размишљају, алфа-таласе замењују асинхрони бета-таласи, који имају мању амплитуду и већу фреквенцију од преко 14 до 30 Хз. Они су повезани са повећаном пажњом и будношћу, а не са перцепцијом. На пример, ако отворимо очи у потпуном мраку и покушамо нешто да видимо, долази до генерисања бета-таласа, иако ништа не опажамо. Региструју се углавном са теменог и чеоног режња приликом њихове активности.
Постоје и гама-таласи, фреквенције од 30 до 80 Хз, који су повезани са вишим менталним функцијама, обједињавањем информација из различитих делова мозга у смисаоне целине, повећаном концентрацијом и будношћу. Осим у будном стању, јављају се и током РЕМ спавања.
Доминантне таласе у биоелектричној активности мозга забележене са спољне површине лобање код здравих одраслих особа, будних, релаксираних и са затвореним очима, чине алфа-таласи, који се претежно региструју са задњих делова мозга, помешани са мањим бројем бета-таласа доминантних у предњим деловима мозга. Одступања од оваквог шаблона која се јаве током будности готово увек су знак болести.
Приликом спавања се ЕЕГ запис значајно мења. Како постајемо поспани, смањује се амплитуда и фреквенција алфа-таласа, а током самог спавања долази до генерисања тета (од 4 до 7 Хз) и делта-таласа (мање од 3,5 Хз), који имају већу амплитуду и мању фреквенцију. Тета-таласи се региструју у теменом и слепоочном (темпоралном) режњу код деце, али су код појединих одраслих особа присутни током емоционалног стреса, посебно разочарања и фрустрације. Делта-таласи често имају два до четири пута већу амплитуду од других можданих таласа, а јављају се само у дубоком спавању и органским болестима мозга. Код новорођенчади су присутни и у будном стању, али с временом ишчезавају. ЕЕГ промене током спавања су праћене променама положаја тела, прага за реаговање на спољашње дражи, лакоћом буђења и бројем покрета тела.
РЕМ и НРЕМ спавање
уредиПостоје, генерално, две врсте спавања, разликовано на основу тога да ли се може видети померање очију иза затворених капака, а то су РЕМ (рапид еyе мовемент; спавање са брзим покретима очију; може), познато и као парадоксално, и НРЕМ, или неРЕМ (нонрапид еyе мовемент; спавање без брзих покрета очију; не може) спавање, које се назива и спороталасно. Таласи током НРЕМ спавања имају велику амплитуду и малу фреквенцију. НРЕМ спавање је подељено у три сукцесивне фазе, где свака има мању фреквенцију и већу амплитуду од претходне. Спавање почиње са проласком кроз све три фазе, што обично траје 30 до 45 минута, а затим се процес обрће и опет се јављају асинхрони таласи мале амплитуде и велике фреквенције, карактеристични за позорно будно стање. Уместо да се пробуди, међутим, особа наставља да спава. Особа која се пробуди за време НРЕМ спавања ретко се сећа да је сањала, али је доказано да се и током НРЕМ спавања сања. Но, док су снови у РЕМ спавању надреални и апстрактни, снови који се јављају у НРЕМ спавању су јасни и сврсисходни. Генерално, НРЕМ спавање се карактерише смањеном метаболичком активношћу мозга, а и проток крви у њему је такође мањи, посебно у можданом стаблу, таламусу, предњем делу хипоталамуса, базалним ганглијама, малом мозгу (церебелуму), као и фронталним, паријеталним и медијалним темпоралним деловима мождане коре.
Друга врста спавања, РЕМ спавање, још се назива и парадоксално јер се особа теже буди, иако је ЕЕГ запис десинхронизован, односно сличан ономе у позорном будном стању. Оно се може описати и као стање менталне будности у телу које спава. Особа која се пробуди за време РЕМ спавања најчешће (у око 80% случајева) се сећа да је сањала, што важи и за људе који се обично не сећају снова када се ујутро спонтано пробуде. Праг буђења у РЕМ спавању је повишен и три пута је виши од оног у дубоком спавању, тј. Н3 фази НРЕМ спавања. Тонус скелетних мишића, који је смањен и током НРЕМ спавања, постаје значајно инхибисан за време РЕМ спавања. Изузети су мишићи ока, па долази до појаве брзих, сакадичних, покрета очију (по чему је овај тип спавања и добио име), дијафрагма и мишићи који подижу и спуштају грудни кош током дисања. РЕМ спавање је много мање окрепљујуће него НРЕМ. Трајање епизода РЕМ спавања се мења, од почетних око 15 минута у првом циклусу на почетку спавања, до 30 и више у следећим, да би пред јутро, тј. буђење, РЕМ фазе биле најдуже. За РЕМ спавање је карактеристично повремено јављање таласа великих амплитуда, који настају у холинергичким неуронима понса и брзо путују до латералног геникулатног једра таламуса, а одатле до окципиталног кортекса. Ови таласи се зову понто-геникуло-окципитални шиљци (ПГО). Понто-геникуло-окципитални таласи се јављају у тзв. фазичном стадијуму РЕМ спавања. Фазични РЕМ стадијум је периодичан и јавља се у склопу тоничког стадијума. Тонички стадијум је заправо РЕМ спавање у ужем смислу јер непрекидно траје током целе РЕМ фазе спавања, а име је добио по атонији великих мишићних група и инхибицији моторике које га карактеришу.[6] До брзих очних покрета и трзања неких мишића долази за време фазичног РЕМ стадијума. Још увек није јасно због чега се НРЕМ спавање периодично прекида РЕМ спавањем, али је примећена улога ацетилхолина у целом процесу. Овај неуротрансмитер има претежно ексцитацијско дејство, везану за изазивање акутне будности и опште ексцитираности нервног система, а уочено је и да лекови са ацетилхолинским деловањем повећавају учесталост јављања РЕМ спавања. Претпоставља се да излазна влакна великих холинергичких неурона у горњем делу можданог стабла могу активисати велике делове мозга, доводећи тако теоријски до типичног стања прекомерне мождане активности, али без буђења, што је карактеристика РЕМ спавања.
За РЕМ спавање је карактеристичан повећан проток крви у мозгу, као и појачана активација неких делова мозга у односу на будно стање. У настанку РЕМ спавања учествују тегментална једра понса, таламус, задњи делови мождане коре, као и лимбички делови мозга, посебно амигдале, које су врло активне за време РЕМ спавања. Са друге стране, фронталне и паријеталне области мозга су већином инхибисане.
Непрекинуто, спавање се одвија циклично, прелазећи из прве и друге у трећу фазу, а затим назад, од треће преко друге, све до 1, где се уместо НРЕМ јавља РЕМ спавање, чиме се завршава један циклус. Обично се током ноћи прође кроз 4 или 5 оваквих циклуса, од којих сваки траје око 90 или 100 минута.[1] Код младих особа на НРЕМ спавање отпада 75 до 80 посто спавања, док остатак припада РЕМ спавању. Епизоде РЕМ спавања трају од 5 до 30 минута, на почетку спавања су краће, а код изразитог умора могу и изостати, док постају све дуже како спавање одмиче.
I поред великог броја истраживања у последњих 60-ак година, као и многих информација о карактеристикама и специфичностима појединих стадијума спавања, сама сврха и функција спавања и даље нису потпуно разјашњене. Док се већина релевантних истраживача слаже како НРЕМ спавање има бар донекле ресторативну функцију, улога РЕМ спавања и даље је предмет расправа и бројних контроверзи.
Полисомнографске карактеристике фаза спавања
уреди
Различите стадијуме спавања први пут су описали Алфред Лумис и његови сарадници 1937. године, када су на основу разлика у ЕЕГ карактеристикама направили скалу од 5 подељака (од А до Е), која је обухватила различита стања, од будности до дубоког спавања.[7] РЕМ спавање је откривено као посебна врста спавања 1953. године, па су Вилијам Демент и Натанијел Клајтман извршили нову класификацију спавања, која је обухватила 4 стадијума НРЕМ и посебну фазу РЕМ спавања.[8]
Алан Рихтшафен и Ентони Кејлс су 1968. године извршили стандардизацију класификације спавања на основу Дементових и Клајтманових резултата и конструисали скалу названу по првим словима њихових имена Р&К Слееп Сцоринг Мануал (у слободном преводу: Р&К приручник за класификацију спавања).[9] По њиховим критеријумима, НРЕМ спавање је такође подељено у 4 стадијума, а 3. и 4. НРЕМ стадијуми су означени као спороталасно спавање (СWС, слоw-wаве слееп). РЕМ спавање је понекад спомињано као пети стадијум.
Америчка академија за медицину спавања је 2004. године извршила ревизију Р&К система класификације. Ревизијом је уведено неколико промена, од којих је најзначајнија подела НРЕМ спавања на 3, уместо на 4 стадијума као раније. Нови резултати објављени су 2007. године, под називом Тхе ААСМ Мануал фор тхе Сцоринг оф Слееп анд Ассоциатед Евентс (Приручник Америчке академије за медицину спавања за класификацију спавања и са њим повезаних појава).[10] Нова класификација обухватила је и стање будности, као и кардиоваскуларне, респирационе и моторне промене.[11][12] НРЕМ спавање је по овој класификацији подељено у 3 стадијума, означених као Н1, Н2 и Н3. Н1 и Н2 стадијуми одговарају првом и другом стадијуму по старој класификацији, док је Н3 обједињени стадијум, који обухвата старе фазе спороталасног спавања, односно трећи и четврти стадијум.
- Током будности ЕЕГ је брз и модулиран, са присутним алфа и бета-активностима; ЕОГ је хаотичан, а ЕМГ показује обиље мишићне активности.
- У првом стадијуму НРЕМ спавања (Н1) ЕЕГ, тј. алфа-таласи, постепено успоравају и до 50%, а јављају се још бета и тета-таласи; ЕОГ показује пендуларне покрете булбуса, док ЕМГ показује оскудну активност. Ово је прелазни стадијум који траје од 1 до 7 минута, из њега се лако буди, а и без буђења се може реаговати на стимулусе, али је упамћивање онемогућено. Уколико се пробуди, особа се често не сећа да је спавала. Ово је први стадијум тзв. површног спавања.
- ЕЕГ код другог стадијума (Н2) показује тета-таласе, са тзв. вретенима спавања, тј. више таласа од 12 до 14 Хз који се јављају 2 до 5 пута у минути, и К-комплексима, спорим двофазним таласима са једним таласом изразито негативне и другим изразито позитивне волтаже, а могу се јавити и делта-таласи; ЕОГ и ЕМГ показују оскудну активност. Ово је најдужи стадијум спавања који траје од 10 до 30 минута, код кога је праг за буђење низак, упамћивање онемогућено, а уколико се пробуди, особа се не сећа шта је сањала.
- Код трећег стадијума (Н3) је присутно више од 50% спороталасне делта-активности, док су ЕОГ и ЕМГ оскудни. Он траје од 10 до 30 минута, а праг за буђење је веома висок. Уколико се особа пробуди, конфузна је и дезоријентисана, нема сећања на снове и понекад аутоматски изводи стереотипне покрете. 3. стадијум се назива и спороталасно спавање, јер је током њега учесталост спорих делта-таласа најинтензивнија.
- Полисомнографија карактеристична за РЕМ спавање подудара се донекле са будним стањем. ЕЕГ је немодулиран, са присутном бета и мањом количином тета-активности; ЕОГ показује брзе покрете, а ЕМГ атонију мишића.
Поред смањеног мишићног тонуса, током спавања долази и до других промена. На пример, за време НРЕМ спавања долази до спорадичног отпуштања гонадотропина и хормона раста из аденохипофизе, смањују се крвни притисак, фреквенција срца, број респирација и интензитет базалног метаболизма за 10 до 30%, а ове најнижу вредност достижу у трећем стадијуму (Н3) НРЕМ спавања. Такође, поменуте физиолошке карактеристике су током НРЕМ спавања врло стабилне. Током РЕМ спавања крвни притисак, срчана фреквенција и дисање нестабилни су и повишени изнад нивоа који имају у будном стању. Повећава се потрошња кисеоника у мозгу и мења терморегулација у складу са спољашњом температуром простора где се спава. Штавише, укупни метаболизам мозга се за време РЕМ спавања може повећати и 20%. Код мушкараца се често, готово увек, јавља ерекција, што је важно за клиничку медицину због утврђивања да ли је еректилна дисфункција (импотенција) физиолошког или психогеног карактера. Најуочљивија карактеристика РЕМ спавања је мишићна атонија. Међутим, иако је мишићни тонус јако смањен, долази до појачане мишићне активности и трзања мишића лица, удова и средњег ува. РЕМ спавање је, дакле, врста спавања при којој је мозак прилично активан, али мождана активност није усмерена тако да особа буде потпуно свесна своје околине, па она истински спава.
-
 ЕЕГ током Н1 стадијума НРЕМ спавања
ЕЕГ током Н1 стадијума НРЕМ спавања -
 ЕЕГ током Н2 стадијума НРЕМ спавања
ЕЕГ током Н2 стадијума НРЕМ спавања -
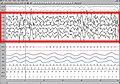 ЕЕГ током Н3 стадијума НРЕМ спавања (стадијуми 3 и 4 по старој класификацији)
ЕЕГ током Н3 стадијума НРЕМ спавања (стадијуми 3 и 4 по старој класификацији) -
 ЕЕГ током РЕМ спавања
ЕЕГ током РЕМ спавања




Теорије, узрасне карактеристике и функција спавања
уредиИако човек проведе близу трећине свог живота спавајући, биолошке функције спавања нису потпуно разјашњене. Спавање има директан и драматичан ефекат само на нервни систем, док је његов утицај на остале функционалне системе мали или чак непостојећи.[1] Особе којима је кичмена мождина пресечена у подручју врата и код којих нема циклуса спавања и будности испод нивоа пресецања не показују штетне ефекте који би могли бити последица непостојања овог циклуса испод тог нивоа.[5] Међутим, да спавање није неопходно, било би нормално да постоје:
- животињске врсте које уопште не спавају,
- животиње које не морају да надокнађују спавање после дужег периода будности и
- животиње на које недостатак спавања нема значајне последице.
Поред неких примитивних животиња које или немају мозак, или је он врло прост, још није откривена животиња која би испунила било који од ових критеријума.[13] Мада одређене врсте ајкула, као што су велика бела или ајкула чекићара, морају бити у сталном покрету како би им се оксидована вода могла кретати кроз шкрге, верује се да оне спавају користећи само једну хемисферу.
Иако су неуронска подручја и трансмитери који се односе на спавање познати, још увек нема објашњења зашто долази до наизменичног смењивања циклуса спавања и будности. Претпоставља се да када нервни центри за спавање нису активни долази до спонтане активације ретикуларних једара мезенцефалона и горњег дела понса. Ова једра изазивају будност (описано у одељку Неурофизиологија циркадијалног ритма и спавања), односно доводе до активације мождане коре и периферног нервног система, који затим шаљу сигнале позитивном повратном спрегом на та иста ретикуларна једра, чиме их још више активишу, па се због те везе будност одржава током дужег периода.
Са друге стране, када мозак остане активан много сати, вероватно долази и до замора неурона самог активацијског система, па нестаје повратна спрега између њих и мождане коре. Тада долази до активације центара за спавање, што омогућава брзи прелазак из стања будности у стање спавања.
Постоји више теорија о спавању, јер оно као феномен фасцинира људе још од периода Античке Грчке. Спомињане су консолидација памћења, конзервација енергије, терморегулација и адаптација на неповољне спољашње услове итд.
Исхемична теорија
уредиИсхемична теорија, на пример, претпоставила је да спавање настаје као последица смањеног дотока крви мозгу или барем центрима за свесност. Старогрчки лекари увидели су везу између каротидне артерије и спавања, а чак је и име ове артерије изведено од грчке речи каро, што значи успавати. Ова тврдња се донекле задржала и у савременој физиологији, јер се сматра да до поспаности која настаје после оброка долази због већег прилива крви из мозга у дигестивне органе. Међутим, ово је једна од првих теорија о настанку спавања, а ако се изузму различита окултистичка веровања, може се сматрати и првом, с обзиром да је ову појаву покушала да објасни на основу објективних чињеница, а не мистичних или магијских феномена. Како је настала у време када нису била доступна сазнања којима савремена физиологија данас располаже, а само на основу посматрања и логичког закључивања, ова теорија се данас узима искључиво као историјска референца.
Ресторативна теорија и ендогени фактори спавања
уредиРазличите теорије о спавању претпостављале су постојање одређених хипнотоксина, односно ендогених фактора и супстанци које изазивају спавање и спровођена су научна истраживања у циљу њиховог изоловања, па је тако утврђено како неки биохемијски агенси у организму заиста имају утицај на појаву спавања. Ресторативна теорија тако претпоставља да је спавање у ствари последица благе интоксикације која настаје због потрошње енергије у будном стању. Према овој теорији, свака контракција мишића или акциони потенцијал доводе до микроскопског цепања ткива. Ово ткиво се затим апсорбује у крвоток и већином се елиминише из организма природним филтерским системима као што су бубрези, плућа, црева или кожа. Међутим, у одређеном тренутку долази до засићења, организам не може да елиминише све продукте разградње и они се накупљају у одређеним деловима мозга, што доводи до измењеног менталног и физичког функционисања. Ове промене се схватају као сигнал да се нагомилани разградни продукти морају елиминисати па добијамо снажну жељу за спавањем. Током спавања су ткива која производе ове материје мање активна па је и разлагање много мање, а долази и до процеса који уклањају већ наталожене разградне продукте из мозга. Другим речима, улога спавања је обнова равнотеже мождане активности.
Заиста, експерименти су показали да цереброспинална течност, крв и мокраћа животиња које су током неколико дана одржаване буднима садрже једну или више супстанци које узрокују спавање ако се другој животињи убризгају у коморски систем. Једна од њих је мурамил пептид, супстанца мале молекулске масе која се накупља у цереброспиналној течности и мокраћи животиња које су биле будне више дана. Само неколико микрограма ове супстанце убризгане у 3. комору изазива после пар минута скоро природно спавање које може трајати више сати. Друга супстанца сличног дејства је нонапептид, који је изолован из крви животиња које спавају, а трећи фактор спавања, који још није идентификован, изолован је из ткива можданог стабла животиња које су више дана одржаване будним. Претпоставља се да продужена будност доводи до накупљања једног или више фактора спавања у можданом стаблу или ликвору, што узрокује спавање.[5] Међутим, с временом је установљено како ове супстанце најчешће не изазивају природно спавање већ неку врсту коме. Ипак, неке од њих заправо изазивају природно спавање, али ниједна сама по себи није нужна и довољна за настанак спавања већ само представљају један од бројних фактора који изазивају његову појаву, па се тако данас једином стварно хипногеном (која изазива спавање) супстанцом сматра аденозин.
Верује се да накупљање аденозина, који између осталог има функцију неуромодулатора, у одређеним деловима предњег мозга смањује будност и индукује спавање. Део стимулативног дејства кофеина потиче од његовог везивања за аденозинске рецепторе и њиховог блокирања. Смањена аденозинска активност подстиче лучење неуротрансмитера допамина и глутамата. Доказано је да примена аденозина или његових агониста изазива дуже спавање, а ниво екстрацелуларног аденозина у неким деловима мозга је већи током будности него за време спавања, и постепено се повећава током дужег периода неспавања. Са друге стране, ниво аденозина за време спавања постепено се смањује.[4]
Еволуциона теорија
уредиЕволуциона или адаптивна теорија спавања говори о еволуционој повезаности периода спавања и будности, тј. активности и неактивности, са преживљавањем у дивљини. Према њеним тврдњама, различите врсте су се адаптирале тако да спавају онда када би будност по њих била најризичнија. Докази су изведени посматрањем животиња. На пример, животиње које имају мање природних непријатеља, као што су лавови или медведи, спавају и по 12 до 15 сати дневно, док оне са више природних непријатеља спавају далеко мање, свега по 4 до 5 сати на дан. Међутим, ова теорија је често оспоравана јер многе животиње не испуњавају њене критеријуме. Спавање се код животиња директно везује за базални метаболизам, па тако пацови, код којих је он врло активан, спавају и по 14 сати дневно, док слонови, чији је базални метаболизам спорији, спавају само 3 до 4 сата на дан. Мада оспоравана, ова теорија је и даље актуелна у проучавању понашања код животиња.
Теорија о конзервацији енергије
уредиОва теорија повезује периоде активности и неактивности са потребама за очувањем енергије. Она спавање људи пореди са хибернацијом, током које се смањују телесна температура и метаболичка активност животиње и тако штеди енергија у одсуству хране. У случају људи, ноћ је непродуктиван период па се током њега спава како би се очувала енергија потребна за дневне активности, обнавља се и повећава ниво гликогена у мозгу који током будности опада. Поред тога, током ноћи је хладније него током дана па би се у случају ноћне активности трошило више енергије за одржавање телесне температуре. Телесна температура подлеже 24-часовним цикличним променама у опсегу од 0,5 до 0,7 °Ц и најнижу вредност достиже током ноћи за време спавања, чиме се смањује губитак топлоте, а постепено се повећава са дневном активношћу. Међутим, многи истраживачи одбацују ову теорију. Поставља се следеће питање: ако је сврха спавања очување енергије, зашто током њега долази до одсуства будности, када се енергија може штедети и у стању мировања? Такође, животиње после буђења из хибернације пролазе кроз период спавања да би надокнадили његов недостатак за време хибернације. Како је током ње дошло до конзервације енергије, очигледно је да то није примарна функција спавања, већ оно постоји због нечег другог.[14] Поред тога, метаболизам можданих ћелија прилично је интензиван и специфичан. Неурони имају минималну способност за анаеробни метаболизам (ослобађање енергије делимичном разградњом гликозе и гликогена без утрошка кисеоника) и скоро сву потребну енергију добијају из гликозе која им се путем капиларне крви допрема из секунде у секунд. Штавише, специфичност неурона огледа се у томе што транспорт гликозе кроз њихову ћелијску мембрану не зависи од инсулина који је неопходан за пренос гликозе у већину других ћелија тела, а количина гликозе која се допреми мозгу за само два минута једнака је укупној количини депонованог гликогена у неуронима.[5] Због тога се поставља питање важности спавања у контексту обнављања нивоа гликогена у мозгу.
Спавање и памћење
уредиДавно је примећена веза између спавања и памћења, и позитиван ефекат који оно има на процес упамћивања нових информација. Претпостављало се да блокирање уласка нових сензорних информација које настаје током спавања омогућава консолидацију памћења. Штавише, истраживања на људима применом позитронске емисионе томографије (ПЕТ) и студије на животињама доказале су да за време спавања може доћи до поновне активације делова мозга који су били активни и приликом упамћивања нових информација, што би могло имати утицаја на консолидацију памћења. Такође, активација ових делова мозга може бити насумична или систематска, чиме би се омогућила интеграција нових информација и оних складиштених у меморији. Међутим, физиолошки механизми овог процеса нису откривени.[15]
Иако су неке од ових теорија добиле мању или већу експерименталну потврду, ниједна није у потпуности одговорила на питање зашто спавамо. Зна се само да током спавања долази до опоравка и одмора делова мозга који су били оптерећени у будном стању и претпоставља се да без њега не можемо живети. Међутим, до сада међу људима није забележен ни један смртни исход као последица физиолошких ефеката недостатка спавања. Када су Вилијама Демента, оснивача Центра за проучавање спавања при Стенфорд универзитету, који се до тада овим истраживањима бавио 50 година, питали зашто спавамо, он је одговорио: По ономе што ја знам, једини одговор на то питање који се може сматрати потпуним је зато што нам се спава.[16]
Мада изостанак спавања у одређеној мери утиче и на друге физиолошке системе, најзначајнији је његов утицај на централни нервни систем. Дуготрајна будност узрок је поремећаја процеса мишљења и може довести до ненормалног понашања, раздражљивости па чак и психотичних епизода, те се стога може претпоставити да спавање обнавља нормалне нивое мождане активности и равнотежу између различитих функција ЦНС. Депривација РЕМ спавања током дужег периода доводи до слабијих психолошких ефеката од депривације НРЕМ спавања.
Утицај форсиране будности на пацове
уредиСтудија из 2007. је утврдила везу између спавања и функционисања имунског система код пацова, јер су пацови који су држани буднима 24 сата имали 20% мање белих крвних ћелија у односу на пацове из контролне групе, чиме је доказано да лишавање спавања нарушава функционисање имунског система. Пацови који су лишавани спавања такође су већ првог или другог дана форсиране будности развијали поремећај терморегулације јер им се унутрашња температура повећала за неколико степени, а константно су губили на телесној маси и поред повећаног уноса хране, повећавала им се срчана фреквенција, развијале се улцерације и крварења у гастроинтестиналном тракту, док је за две до три недеље долазило до смртног исхода. Сличне последице су забележене и код пацова који су били лишавани само РЕМ спавања, с том разликом што су се патолошки процеси развијали спорије, а до смртног исхода је долазило за четири до пет недеља.[15] Испитивања и на другим животињама дала су сличне резултате са крајњим смртним исходом код паса, зечева, у нешто мањој мери код мачака, чак и код воћних мушица.
Међутим, упркос већем броју истраживања, још увек није јасно зашто је недостатак спавања довео до смрти код експерименталних животиња, али су на основу добијених резултата научници претпоставили да спавање, макар код (неких) животиња, има бар једну виталну функцију.[15]
Експериментално је доказано да спавање утиче на зарастање рана код животиња и студија из 2004. године показала је да депривација спавања успорава зацељивање опекотина код пацова.
Спавање и развој
уредиПостоје истраживања о онтогенетској улози спавања и тврдње да РЕМ спавање има улогу у развоју мозга. Резултати Марксовог истраживања из 1995. године указују на то да недостатак РЕМ спавања у раном животу може довести до каснијих бихевиоралних сметњи, поремећаја спавања, смањене мождане масе и патолошке апоптозе неурона. Код беба, највећи део спавања чини управо РЕМ спавање, док превремено рођени младунци неких животиња више времена проводе у РЕМ спавању. Истраживачи такође тврде да инхибиција мишићног тонуса при РЕМ спавању постоји да би се спречиле евентуалне повреде детета које би могле настати због неконтролисаних моторних радњи до којих би дошло да тонус није инхибисан, па појачана мождана активност нормална за РЕМ спавање може да се одвија без бојазни од ових повреда и омогући тако формирање и матурацију синапси. Постоје студије о вези између недовољног спавања и менталних болести.
Спавање и ментални процеси; мањак спавања
уреди.jpg)
НРЕМ спавање се сматра неопходним за инхибицију деловања неуротрансмитера, дозвољавајући рецепторима да се одморе и не развију толеранцију на ове хемијске супстанце, па тако моноамини (серотонин, норадреналин и хистамин) могу испољавати своја уобичајена дејства током будности. То утиче на расположење и способност учења. Компромитовано РЕМ спавање јавља се у склопу клиничке депресије, за шта се претпоставља да је последица поремећаја холинергичког система мозга, на пример, претеране осељивости на ефекте ацетилхолина. Поремећаји РЕМ спавања код депресије обухватају скраћено време (латенцу) између два РЕМ циклуса и дуже трајање РЕМ спавања. Лекови који смањују РЕМ спавање, као што су антидепресиви, ефикасни су у лечењу симптома депресије, док лекови који продужавају РЕМ спавање изазивају или погоршавају ове симптоме.[2]
Дуготрајна будност праћена је дужим периодом или већим интензитетом спавања, или и једним и другим. Смањује се латенца успављивања, тј. време потребно за прелазак из стања будности у стање спавања. Повећава се трајање НРЕМ спавања (посебно стадијума 3 и 4, односно Н3), као и његов интензитет. Такође се повећава и трајање РЕМ спавања, али је непознато да ли му се повећава и интензитет.[15]
Многи људи, као што су лекари, медицинске сестре, полицајци итд., имају лошу хигијену спавања, тј. поремећен циклус спавања услед сменског карактера посла којим се баве. Они често показују симптоме недостатка спавања[17]. Једно америчко истраживање на узорку здравих људи старости између 50 и 65 година показало је да се просечно време спавања између 1959. и 1980. године смањило за један сат, а као узроци се између осталог наводе повећана индустријализација, све већи број људи који раде по сменама, развој телевизије.[6]
Дуготрајно неспавање има највећи утицај на расположење, али и моторне као и когнитивне функције, посебно пажњу и радну меморију, што може довести до фаталних последица, на пример саобраћајних несрећа или повреда при управљању индустријским машинама. Како су нарушени радна меморија и обрада информација, вид, продужено време реакције и расуђивање, уз честу појаву халуцинација, врло лако долази до различитих незгода. Међутим, за њих је често одговоран још један феномен који се зове микроспавање. Ово је, у ствари, епизода спавања која наступа после периода дуже будности и може трајати мање од секунде па све до пола минута. Наступа без упозорења, из стања будности се нагло прелази у први стадијум НРЕМ спавања, а људи су га несвесни, описујући га као осећај вртоглавице, не знајући да су заправо накратко заспали. Појава микроспавања повезана је са трајањем будности, па је тако његово јављање после 40 сати неспавања готово извесно. Такође се нарушава концентрација, посебно на више циљеве или идеја истовремено, особа постаје мање спонтана и више ригидна, смањује јој се жеља за дружењем, говор постаје неразговетнији и отежан, јављају се збуњеност и дезоријентација, неспретност, лошија обрада информација и функционисање краткорочне меморије, неодлучност.
Интензитет периферног метаболизма се код људи и животиња повећава ако су дуже време будни (код људи и у случају инсомније). Са друге стране, укупни метаболизам мозга се смањује у односу на онај у будном стању при нормалним околностима, посебно у таламусу и међумозгу. Утврђено је да дуготрајна будност доводи до смањене толеранције на гликозу, повећане активности симпатичког нервног система и појачане секреције кортизола, што указује да би неспавање могло утицати на појаву дијабетеса, хипертензије и гојазности. Такође, пацијенти са инсомнијом често обољевају од других болести, посебно кардиоваскуларних, што може бити потврда за повезаност неспавања са утицајем на здравље. Међутим, јасна веза још увек није утврђена, а форсирана будност код људи у експерименталне сврхе се углавном не спроводи дуже од три до четири дана (званично најдужи период износи 11 дана), па је за сада немогуће установити да ли дуже неспавање може имати штетније последице по здравље.[15] Без обзира што неиспавану особу можемо лако препознати по спољном изгледу и променама у понашању, неуролошке промене су релативно мале и лако реверзибилне. У истраживању ефеката неспавања током периода од неколико дана примећена је појава благог нистагмуса (невољна ритмичка осцилација очних јабучица), тремора руку, неартикулисаног и неразговетног говора и птозе (спуштања горњих очних капака). После дужег периода будности уочавају се и снижен корнеални рефлекс, појачани фарингеални рефлекс, мишићни (дубоки) рефлекси, убрзано лупање срца, већа резистенција на инсулин, глад, као и појачан сензибилитет за бол. Међутим, сви симптоми нестају непосредно после надокнађеног спавања. Такође, долази и до значајних промена ЕЕГ активности. У великом броју истраживања забележено је прогресивно смањивање алфа-таласа током форсиране будности. Епизоде алфа-таласа код испитаника после 24 сата будности трају до 10 секунди, смањујући се на 4 до 6 секунди после 72 сата, односно на 1 до 3 секунде после 120 сати. Након 115 сати будности, затварањем очију престају да се генеришу алфа-таласи. Проценат делта и тета-таласа повећао се са 17% и 12% на 38% односно 26%. Појачана делта-активност је била најуочљивија у предњим деловима мозга младих људи.[18]
Дужи недостатак спавања чини особу подложнијом утицајима и сугестијама других људи, па се форсирана будност често користи као облик мучења или изнуђивања исказа.
Генерално, сматра се како НРЕМ спавање има битну улогу у ресторативном процесу обнављања природне равнотеже између нервних центара, док физиолошка функција РЕМ спавања остаје непознаница, а истраживања показују како изостанак ове фазе спавања код људи чак и у периоду од две недеље не изазива озбиљне или чак икакве последице по понашање.
Уопштено говорећи, нису сви људи и све активности подједнако погођене недостатком спавања. Психолошки ефекти биће израженији и озбиљнији код особа са менталним поремећајима. Такође, истраживања показују како мањи недостатак спавања не утиче негативно на физичке активности и вежбе које не захтевају већи ментални напор, као што су трчање, дизање тегова и пливање, с тим што се у њихово обављање улаже већи напор и потребно је дуже времена за одмор. Са друге стране, неспавање значајно утиче на постигнуће у спортовима који захтевају више пажње, размишљања и координације, као што су фудбал, кошарка и тенис. Задаци који захтевају логичко размишљање су успорени, али нису угрожени.
Случај Рендија Гарднера
уреди1963. године, Ренди Гарднер, тада 17-годишњи средњошколац, извео је специфичан експеримент за научну приредбу у Сан Дијегу.[4] Почео је 28. децембра у 6 сати ујутро, да би после 11 дана (264 сата) постао светски рекордер у најдужем временском периоду проведеном без спавања. Притом није користио никакве лекове, чак ни кофеин. Било је то тешко искуство. После два дана неспавања, Ренди је постао раздражљив, било му је мука, имао је проблема са упамћивањем и концентрацијом. Четвртог дана је био изразито слаб и имао је мање делузије (сумануте идеје) , док се седмог дана јавио и тремор, неразговетан говор, а у ЕЕГ запису више није било алфа-таласа. Једанаестог дана, када је од њега тражено да, почевши од броја 100, броји уназад, одузимајући притом 7, стао је на броју 65. Када је упитан зашто је стао, одговорио је како је заборавио шта је радио.[19]
Међутим, Ренди, упркос предвиђањима појединих стручњака, није постао психотичан, а чак је и на конференцији за штампу која је организована после обарања рекорда говорио разговетно и разложно. Када је после овога отишао на спавање, спавао је скоро 15 сати, да би после тога провео још 23 сата будан, чекајући ноћ, а затим је спавао још 10 и по сати. После првог периода спавања од 15 сати, већина симптома је нестала, док му се циклус спавања у року од недељу дана вратио у пређашње стање. Најинтересантнија чињеница у вези са Рендијевим експериментом је то што није било трајних последица по њега, за разлику од пацова, који после одређеног периода форсиране будности умиру.[4] Гарднер је и данас званични рекордер, мада постоје тврдње о обарању његовог рекорда, али ти случајеви нису праћени и регистровани.
Фатална фамилијарна инсомнија
уредиМада није јасно утврђена повезаност недостатка спавања и озбиљнијих физиолошких дисфункција, постоји врло ретка болест која онемогућава спавање. То је фатална фамилијарна инсомнија (смртоносна породична несаница). Ово је прионска болест мозга која се налеђује аутозомно-доминантно: ако је један родитељ носилац мутације, проценат обољевања потомства је 50%. Прионски протеини налазе се на ћелијским мембранама, претежно неуронским, а када дође до трансформације нормалног (ПрПC) у патолошки (ПрПСц) облик овог протеина, долази до неке од прионских болести, од којих је вероватно најпознатија говеђа спонгиформна енцефалопатија (болест лудих крава). Све ове болести су неизлечиве, а карактеришу их спонгиформна (сунђераста) дегенерација (отуд и назив спонгиформне енцефалопатије), губитак неурона мозга и пролиферација астроцита.[1][20][20]
Фатална фамилијарна инсомнија настаје због мутације гена на кратком краку хромозома 20, који доводи до трансформације нормалног облика прионског протеина у патолошки.[21] На свету постоји само 40 породица у оквиру којих има носилаца ових гена. Болест се јавља у различитим узрастима, углавном између 18. и 60. године, просечно у 50. години живота.[22] Симптоми често варирају, чак и међу случајевима у истој породици. Најпре се јавља прогресивна несаница, а затим и напади панике, фобије и параноја. После долази до халуцинација, потпуне немогућности спавања и губитка телесне масе. На крају се јавља деменција, када пацијент престаје да реагује на спољашње дражи и долази до дефицита говора. Ово је последњи стадијум болести, после кога наступа смрт. Смртни исход обично наступа у року од неколико месеци, до две или три године. Код фаталне фамилијарне инсомније са краћом клиничком сликом (где смрт наступа за мање од годину дана), од почетка долази до готово потпуне несанице, а спонгиформне дегенерације су углавном ограничене на таламус и доња оливарна једра и не шире се на кортекс. У типу болести са дужом клиничком сликом (где смрт наступа за 2 до 3 године), несаница се постепено развија, а спонгиформне дегенерације захватају велике делове мождане коре. Иако многи симптоми ове болести настају због спонгиформних дегенерација, верује се да и недостатак спавања сам за себе утиче на ток болести, посебно код облика са краћом клиничком сликом, јер компликације и тежина болести корелирају са тежином инсомније, а не са нагомилавањем прионских протеина.[15]
Не постоји лек за фаталну фамилијарну инсомнију. Барбитурати и хипнотици нису дали ефекта у лечењу, а чак могу и погоршати клиничку слику. Било је и покушаја да се код пацијента седативима индукује кома, како би се успоставио циклус спавања, али је ЕЕГ запис и после тога показивао мождану активност карактеристичну за будно стање.[23]
Узрасне карактеристике и количина спавања
уредиКарактеристике спавања се с годинама мењају, пратећи развој и матурацију нервног система, од оних карактеристичних за новорођенче, до типичног спавања одраслих особа. Спавање има своју структуру која је квалитативно јединствена код здравих људи, али се по квантитету разликује у појединим узрасним групама. Јасан циклус спавања и будности успоставља се око 24. недеље трудноће, док се разликовање неке врсте "активног" (РЕМ) и "тихог" (НРЕМ) спавања јавља у последњем тромесечју. Структура и дужина спавања код здравих особа зависи од узраста и јасан циклус НРЕМ-РЕМ спавања се успоставља око 16. недеље живота, али траје краће него код старије деце и с годинама се продужава. Иако постоје индивидуалне варијације, одрасли просечно спавају једнократно ноћу 7 до 8 сати. Новорођенчад спавају у више наврата (вишефазно) и до 18 сати на дан, где је заступљеност РЕМ спавања око 50%, (код недоношчади и до 80%) али се број периода спавања и његова дужина прогресивно смањују и достижу вредност као код одраслих између 4. и 10 године живота. Тако заступљеност РЕМ спавања опада на 30 до 35% између 9. и 12. месеца, да би код људи између 50 и 70 година старости чинило само око 15% целокупног спавања .[1][6][24] Генерално, код новорођенчади у другој недељи живота је дневно спавање једнако ноћном и износи око 8 сати, тј. укупно око 16 сати, да би већ у 16. недељи беба дању спавала 4 до 5, а ноћу 9 до 10 сати, а циркадијални ритам се даљим сазревањем постепено стабилизује. Деца предшколског узраста обично и даље спавају преко дана, али се са овом праксом најчешће прекида када редовно крену у школу. Код њих се јавља повишен проценат спороталасног спавања са високим прагом за буђење. Међутим, током адолесценције се смањује учесталост спороталасног спавања, верује се због губитка синапси, а адолесценти и генерално мање спавају, али се то више тумачи променама у понашању него смањеном потребом за спавањем.
Стари људи спавају у просеку 4 до 6 сати, односно, спавање се код њих скраћује и постаје лакше. Међутим, сматра се да код старих људи не долази до значајније промене у потреби за спавањем у односу на потребе здравих људи средње доби, већ они мањак спавања током ноћи надокнађују поподневним одмором, тзв. дремањем. Такође, постоје и индиције да је спавање краће и слабије код старијих људи због смањене количине мелатонина који лучи епифиза. Са годинама наставља да се смањује удео спороталасног спавања у целокупном циклусу, које до 60. године чак може и потпуно нестати, а спавање постаје испрекидано, са повећаном латенцом успављивања, више буђења током ноћи и више спавања током дана. Старији људи се често буде раније. Проценат РЕМ спавања се не смањује или се смањује тек незнатно код здравих људи, али код људи болесних од Алцхајмерове и других неуродегенеративних болести које погађају централни нервни систем оно може потпуно изостати.
Трајање спавања одређује ДЕЦ2 ген. Људи који су носиоци мутације овог гена спавају два сата мање него нормално. Професорка неурологије Јинг-Хуи Фу и њене колеге у лабораторији узгајају мишеве носиоце мутације ДЕЦ2 гена, који спавају мање од нормалних мишева.[25][26]
Неки научници деле људе на вечерње (сове) и јутарње типове (шеве).[6] Јутарњи типови лако се буде и врло су продуктивни ујутро. Умарају се и одлазе на спавање рано увече. Са друге стране, вечерњи типови се тешко буде ујутро, а када се пробуде осећају се уморно. Постају одморнији како се приближава вече. Најпродуктивнији су увече, касно лежу и устају касније ујутро. Истраживања су показала да вечерњи и јутарњи типови имају различите механизме терморегулације, где код јутарњих типова телесна температура достиже дневни максимум сат времена раније у односу на вечерње.
Различити спољашњи и унутрашњи фактори утичу на потребу за спавањем, па тако физички рад, болести, психолошки стрес, повећања ментална активност и трудноћа доводе до повећања жеље за спавањем.[2]
| Узраст или физиолошко стање | Просечна дневна количина спавања |
|---|---|
| Новорођенчад | до 18 сати |
| Бебе од 1 до 12 месеци | 14 до 18 сати |
| Деца од 1 до 3 године | 12 до 15 сати |
| Деца од 3 до 5 година | 11 до 13 сати |
| Деца од 5 до 12 година | 9 до 11 сати |
| Адолесценти | 9 до 10 сати |
| Одрасли људи средње доби | 7 до 8 сати |
| Стари људи | 4 до 6 сати |
| Труднице | 8(+) сати |
Табела 1: Разлике у времену спавања за одређене узрасне групе и физиолошка стања
Снови
уреди
Сан је ментална активност која се јавља за време спавања. То је невољан и ирационалан мултисензорни доживљај сличан халуцинацији, који не подлеже критичности, а онај који сања у њему се обично појављује као активни учесник, а не као посматрач. Снови се везују за РЕМ спавање, али се могу јавити и током спороталасног спавања, но, они се углавном заборављају, односно не долази до њихове консолидације у памћењу.
Сновима је вероватно највећа пажња посвећена у оквиру психоанализе, која је истраживала несвесне садржаје сматране за узрочнике неуроза. Сигмунд Фројд је у својој књизи Тумачење снова објављеној 1900. године назвао снове царским путем ка несвесном и ту је опширно писао о сложеним везама између свесног и несвесног психичког живота. По његовој теорији, осећања која су психолошки потиснута у несвесном делу ума често се манифестују у сновима или се испољавају као душевне и психосоматске болести. Према Фројду, највећи део потиснутих осећања чине скривене и забрањене сексуалне жеље.[6] Фројд на снове гледа крајње позитивно, сматрајући их својеврсним заштитним вентилом који чува душевно здравље. С временом је цео психоаналитички концепт напредовао и био дорађиван, па у завршној фази формирања психоаналитичких идеја несвесно није обухватало само друштвено неприхватљиве жеље, већ је цео психички живот претежно схваћен као несвестан, са незнатним свесним делом. Фројд је 1923. године, у 67. години, формулисао своју познату структуралну теорију личности, по којој су основне структуралне јединице личности Ид, Его и Суперего. Ид је схваћен као нагонски део личности који садржи подстицај за непосредно задовољење потреба по сваку цену и без одлагања, без уважавања моралних забрана. Суперего представља друштвену компоненту личности и садржи идеале којима тежимо; циљ му је успостављање контроле понашања у складу са правилима друштва, али често функционише неусклађен са реалношћу. Его се руководи принципом реалности и његова примарна улога је да задовољи жеље проистекле из Ида, али уз поштовање моралних принципа из Суперега.[27] По Фројду, снови настају због губитка контроле свесног Ега над несвесним Идом.
Модерна наука сматра Фројдову теорију застарелом већ неколико деценија и тежи ка објашњавању феномена снова на основу физиолошких и анатомских карактеристика. Међутим, Фројд неоспорно мора бити споменут због великог утицаја који је његова психоаналитичка теорија личности имала не само на психологију и психијатрију, већ и на медицину уопште, филозофију и уметност, посебно с обзиром на прилично скромна знања о неуробиологији која су у то време била доступна.
Једна претежно психолошка теорија сматра како је у сновима заступљено понашање које се иначе ретко јавља у понашању током будног стања. Иако њене поставке нису потврђене нити оспорене, истраживања су показала како око 60% садржаја снова чине туга, страх или бес; 20% срећа и радост, а само 10% се односи на садржаје повезане са сексуалним радњама, што је прилично изненадило истраживаче.[28]
Једно другачије схватање сматра да је еволутивна функција снова филтрирање непотребних сећања која се нагомилавају током дневних активности. Франсис Крик тако сматра да су снови механизам за елиминацију паразитских садржаја који би у супротном превагнули и довели до поремећаја процеса мишљења.[28]
Истраживање снова добило је нови замах када је 1953. године откривено РЕМ спавање. Психоанализа и психодинамика су дотад била основни концепти научне психијатрије. Претпоставка је да се 80% сањања одвија током РЕМ, а само 20% за време НРЕМ спавања. РЕМ-снови се много лакше памте, најчешће су живописни, емотивно обојени, нереални, сложени и бизарни. Са друге стране, некад је могуће делимично упамћивање снова после буђења из НРЕМ спавања, а ти снови су углавном реалистичнији. Класична и интракранијална ЕЕГ испитивања (иЕЕГ), као и магнетоенцефалографске (МЕГ) студије показују како за време РЕМ спавања долази до значајно већег генерисања гама-таласа у односу на НРЕМ спавање. Како су гама-таласи у будном стању повезани са повећаном менталном активношћу и концентрацијом, сматра се да њихово појачано јављање у РЕМ фази спавања има везе са когнитивним, меморијским и другим процесима који заједно утичу на целокупни (претежно визуелни) доживљај самог сна.
Даљи напредак технологије омогућио је примену снимања функционалном магнетном резонанцијом (фМРИ) и ПЕТ скенерима у научним истраживањима, самим тим и детаљније упоређивање мождане активности у будном стању и током РЕМ спавања, као и боље разумевање свести уопште. Активност у амигдалама, тегменталним једрима понса и предњем делу цингуларне коре се повећава током РЕМ спавања, док се активност у задњем делу цингуларне коре и префронталном кортексу смањује. Повећана активност у лимбичком систему током РЕМ фазе, а смањена у префронталном кортексу може објаснити одређене карактеристике снова (емотивну обојеност, ирационалност и слично).[28]
Снови нису црно-бели, већ сањамо у боји. Током њих се могу ангажовати свих пет главних чула, од којих је најдоминантније чуло вида, а за њим следи чуло слуха. Чула додира, мириса и укуса много су мање заступљена. Снови могу бити пријатни или непријатни, срећни или тужни..., а често обухватају жеље, страхове и друге делове из живота сањача. Смисао снова је ретко рационална, већ се они одвијају у маниру хаотичног смењивања ликова, догађаја и места, са често бизарним и надреалним мешањем ових елемената.
Да су снови повезани са нашим физиолошким стањем и окружењем током будности говори и чињеница да људи који су слепи од рођења немају визуелне снове, већ само у аудитивном или неком другом модалитету који не захтева виђење. Ако услед можданог удара или друге врсте оштећења ткива човек изгуби способност опажања боја, снови постају ахроматски. Такође, деца ретко пријављују снове пре 4. или 5. године, а и у том случају они врло кратко трају. То је у складу са чињеницом да деца тог узраста имају ограничену способност имагинације и нису у могућности да вербализују сложене и емотивно обојене исказе.[15]
Неуробиолошка функција и значај снова и даље су непознаница. Према теорији активације и синтезе, коју су предложили професори са Харварда Алан Хобсон и Роберт Макарли, током РЕМ спавања долази до повећане активности неурона у различитим деловима мозга, највише у можданом стаблу, одакле се она даље преноси на друге делове мозга, првенствено базалне (лимбичке) регионе, који се сматрају повезаним са емоционалним доживљајима, а затим и поједине делове мождане коре, где долази до побуђивања различитих сензорних зона, што, претпоставља се, доводи до интеграције ових сигнала и изазивања доживљаја снова. По њима, ПГО таласи (види пододељак РЕМ и НРЕМ спавање) из понса који преко латералног геникулатног једра таламуса долазе до видне коре бивају протумачени као видне информације, што доводи до визуелног доживљаја снова. Међутим, ова теорија не објашњава како такви насумични нервни импулси доводе до сложених и повезаних прича које се често одвијају у сновима, или зашто се неки снови понављају.[4]
Погрешно је схватање снова и РЕМ спавања као исте појаве, или поистовећивање снова и спавања уопште, што је честа појава међу лаицима. Већина истраживања везаних за спавање снове испитује индиректно, на основу РЕМ спавања, јер се они не могу објективно мерити и посматрати, док РЕМ спавање може. С обзиром на немогућност испитивања непосредних карактеристика снова, многи научници се њима директно и не баве у склопу изучавања карактеристика спавања.
Неки истраживачи претпостављају да су снови кључни за консолидацију памћења и да за време сањања долази до филтрирања памћења, када се губе непотребне и несврсисходне информације.
Међутим, и поред више теорија и бројних научноистраживачких напора, објашњења функције снова и сањања и даље су хипотетичка.
Поремећаји спавања
уредиСтални проблеми са спавањем су релативно учестала појава код људи, што је чест разлог значајних сметњи у њиховом свакодневном функционисању или погоршања других постојећих болести.[1] Сматра се да 40% популације САД у неком периоду живота има проблема са одређеним поремећајем спавања. Учесталост ових поремећаја се повећава са годинама и више се јављају код жена него код мушкараца.[28]
Болесници могу да се жале на неколико врста тегоба везаних за поремећаје спавања и будности. Примарни поремећаји спавања подељени су у две групе: дисомније и парасомније. Код дисомнија је компромитовано време проведено у спавању и оне се деле на инсомније и хиперсомније. Парасомније се односе на абнормална понашања током спавања или у периоду који води од стања спавања до стања будности, или на појаву нормалног спавања у неадекватно време.[2]
Несаница (инсомнија)
уреди
Несаница је поремећај који карактерише недовољан квантитет и/или квалитет спавања. Барем 50% људи доживи повремену несаницу, док се процењује да 15 до 20% има хроничну несаницу, која доводи до озбиљних поремећаја у дневним животним активностима, као и до повећаног ризика од повреда несрећним случајем. Такође, она може погоршати друге соматске болести, а највећи проблем је што већина случајева остаје недијагностикована.
Несаница изазвана егзогеним утицајем
уредиОва врста несанице није дуготрајна и најбоље се лечи, када је то могуће, отклањањем чиниоца који је изазива. Најчешћа несаница овог типа је пролазна ситуациона инсомнија која се може јавити при промени амбијента спавања (одлазак на пут, ноћење у хотелу, шатору, боравак у болници...). Други важан чинилац који је може изазвати је психолошки или физиолошки стрес. До опоравка долази за неколико дана, а у неким случајевима се може лечити применом хипнотика. Међутим, узимање хипнотика у случају ове врсте несанице треба да траје што краће.
Несаница удружена са психијатријским болестима
уредиНесаница се јавља код 80% психијатријских поремећаја, најчешће код депресије и посттрауматског стресног поремећаја, и често је први знак психијатријске болести. Чак и када је невезана за психијатријски поремећај, дуготрајна несаница повећава ризик од појаве депресије. Уз терапију основне болести, несаница се лечи хипнотицима.
Синдром немирних ногу и несаница
уредиСиндром немирних ногу се карактерише осећајем надолажења пузеће непријатности у дубини потколеница и бутина, што доводи до неодложиве потребе за померањем ногу, које само привремено отклања непријатност. Осећај се јавља сваки пут када је особа у стању мировања пред уснивање. Померање ногу ремети заспивање и изазива тешку несаницу и поред велике жеље и потребе за спавањем. Поремећај траје целог живота, а последице су често драматичне, уз наглашене брачне проблеме јер супружници не спавају у истом кревету. Лечење се доста успешно спроводи применом малих доза леводопе.
Прекомерна дневна поспаност (хиперсомнија)
уреди
Хиперсомнија у великој мери смањује радну и животну способност, а најчешћи узроци овог поремећаја су апнеја у спавању и нарколепсија. Такође, злоупотреба наркотика и одређени лекови могу изазвати пролазну хиперсомнију.
Синдром апнеје у спавању
уредиПрекиди дисања за време спавања су чест и озбиљан узрок претеране дневне поспаности и поремећеног спавања. Прекид дисања доводи до учесталог кратког буђења кога болесник не мора бити свестан, скраћује се или потпуно изостаје НРЕМ фаза спавања, а последице су онеспособљавајуће нарушавање будности током дана, повећан ризик од изазивања саобраћајних удеса, хипертензија и друге озбиљне кардиоваскуларне компликације, затим повећан морталитет. Прекид дисања настаје или због оклузије ваздушног пута (опструктивна апнеја у спавању), недостатка респираторног напора (централна апнеја у спавању) или због комбинације оба фактора (мешовита апнеја у спавању). Апнеја се чешће јавља код гојазних људи, а једини довољно ефикасан начин лечења је коришћење апарата који преко посебне маске, која се носи при сваком спавању, удувава ваздух под повишеним притиском у плућа (ЦПАП).
Нарколепсија
уредиНарколепсија је болест у којој доминирају поремећај способности да се вољно одржи будност и поремећај регулације РЕМ спавања, а тежина варира ретких и безначајних сметњи до тешке онеспособљености. Ова болест се карактерише наступањем спавања на које особа не може да утиче, у ситуацијама када се оно нормално не јавља. Клиничка слика ове болести састоји се од четири фактора: 1. претеране дневне поспаности, 2. губитка мишићног тонуса изазваног јаким емоцијама, уз очување свести (катаплексија), 3. халуцинација на почетку спавања (хипнагогних халуцинација) или при буђењу (хипнопомпних халуцинација) и 4. парализе мишића приликом уснивања или буђења (парализа спавања). Верује се како је поремећај повезан са абнормалностима у функционисању холинергичких неурона.
Парасомније
уредиПарасомније су абнормална понашања која настају због или се дешавају за време спавања. Парасомније настају из НРЕМ спавања, има их више врста и ту се убрајају месечарење (сомнабулизам), ноћни страхови (павор ноцтурнус), ноћно умокравање (енеуресис ноцтурна), шкргутање зубима (бруxисмус), причање током спавања, пароксизмална ноћна дистонија и ноћни грчеви, ноћно трешење главе исл. Месечарење се чешће јавља код деце него код одраслих, а претежно погађа мушкарце. Епизода месечарења обично траје неколико минута. Месечари ходају отворених очију, избегавајући физичке препреке у окружењу, али се при буђењу не сећају ових догађаја.
Спавање код животиња
уреди
Проучавање спавања код животиња важно је за разумевање спавања људи. Чињеница да је оно међу животињама врло распрострањено, чак и у условима када је то опасно, уакзује на то да спавање вероватно има битну биолошку функцију. Томе доприносе и примери неких задивљујуће развијених адаптивних процеса, као што је једнохемисферно спавање.[15] Верује се како се спавање јавља код свих кичмењака.[29]
Од свих животиња, спавање је највише проучавано код сисара, а НРЕМ и РЕМ спавање су забележени код свих сисара и торбара. Чак и фосоријални сисари, на пример кртице, чији је видни систем слабије развијен, показују периодичну ЕЕГ активност сличну РЕМ спавању.[15]
Код неких животиња је тешко забележити неурофизиолошке промене које настају за време спавања, па се оно изучава на основу бихевиоралних карактеристика, као што су мимика, положај тела и смањено реаговање на спољашње дражи. Из спавања се лако буди, за разлику од хибернације или коме, а епизода спавања која настаје после пролонгиране будности дуже траје и спавање је дубље. Биљоједи, којима је потребан већи период будности за налажење хране, обично током дана спавају мање него месождери сличне величине, који могу одједном појести довољно меса за неколико дана.
Коњи и други унгулатни биљоједи могу спавати стојећи, али такође морају лећи током краћег периода РЕМ спавања, које изазива атонију мишића. Слепи мишеви спавају висећи наглавачке. Код неких водених сисара и птица током спавања једна половина мозга може бити активна, што се зове једнохемисферно спороталасно спавање.[30] Ово је карактеристично и за делфине и китове, због потребе да се одржи дисање: они морају с времена на време изронити на површину да узму ваздух. Активност и неактивност наизменично се смењују између две хемисфере. Тако кљунасти делфини током 12-часовног циклуса спавања 2 сата спавају једном хемисфером, затим проводе 1 сат будни, да би после тога спавали 2 сата другом хемисфером.[4]
Осим код сисара, смењивање НРЕМ и РЕМ спавања пронађено је једино код птица. Мада проводе приближно исту количину времена спавајући као и сисари, РЕМ фаза код птица је за четвртину краћа. Такође су присутни атонија мишића и промене у терморегулацији. Постоје тврдње да неке птице спавају у лету, али то није потврђено. Птице могу, као и водени сисари, држати једно око отворено током НРЕМ спавања, што је повезано са појачаном ЕЕГ активацијом у супротној хемисфери мозга (у односу на страну на којој је отворено око). Време проведено спавајући са отвореним оком повећава се уколико је ситуација ризичнија, на пример, када се птица налази на ивици јата. Штавише, отворено око је тада усмерено не ка другим птицама, већ у даљину, као да пази на грабљивице.[15]
Младунци многих сисара у првим месецима живота спавају дуго, током већег дела дана.[31] Међутим, китови убице и неки делфини уопште не спавају током првих месеци живота.[32] То се објашњава способношћу копнених сисара да лако заштите своје младунце док спавају, док морске животиње, чак и као врло младе, морају константно бити на опрезу због грабљиваца.
Донедавно је постојао мали број истраживања и података везаних за спавање бескичмењака, као што су инсекти. С обзиром на слабу могућност бележења мождане активности код инсеката, научници су се претежно фокусирали на уочавање понашања које би могло бити повезано са спавањем. Овакво понашање уочено је код неколико врста инсеката, док је мождана активност карактеристична за спавање забележена једино код пчела и воћних мушица. Присутна је мања мождана активност током спавања, док су неурони видног система пчела мање осетљиви на визуелне стимулусе за време спавања него током будности. Такође, као и код сисара, време спавања се и у воћних мушица смањује са годинама, а показују и исту реакцију на деловање неких хемијских агенаса, на пример кофеина.[15]
Литература
уреди- Демент, Wиллиам. Тхе Еффецт оф Дреам Деприватион. Америцан Ассоциатион фор тхе Адванцемент оф Сциенце.
Види још
уредиРеференце
уреди- ^ а б в г д ђ е ж з Костић, V. (2007). Неурологија за студенте медицине. Београд: Медицински факултет.
- ^ а б в г Садоцк, Б., Алцотт Садоцк, V. (2007). Каплан & Садоцк'с Сyнопсис оф Псyцхиатрy: Бехавиорал Сциенцес/Цлиницал Псyцхиатрy (Тентх Едитион). Пхиладелпхиа: Липпинцотт Wиллиамс & Wилкинс.
- ^ Барретт, К., Барман, С., Боитано, С., Броокс, Х. (2010). Ганонг'с Ревиеw оф Медицал Пхyсиологy (Тwентy-Тхирд Едитион). Неw Yорк Цитy: Тхе МцГраw-Хилл Цомпаниес, Инц.
- ^ а б в г д ђ Беар M., Цоннерс, Б., Парадисо, M. (2007). Неуросциенце: Еxплоринг тхе Браин (Тхирд Едитион). Балтиморе: Липпинцотт Wиллиамс & Wилкинс.
- ^ а б в г Гуyтон, А., Халл, Ј. (2008). Теxтбоок оф Медицал Пхyсиологy (Елевентх Едитион) [Медицинска физиологија]. Београд: Савремена администрација.
- ^ а б в г д Судхансу, C. (2009). Слееп Дисордер Медицине: Басиц Сциенце, Тецхницал Цонсидератионс, анд Цлиницал Аспецтс (Тхирд Едитион). Пхиладелпхиа: Саундерс Елсевиер.
- ^ Лоомис, Алфред L; Харвеy, Е. Н.; Хобарт ГА (1937). „ИИИ Церебрал статес дуринг слееп, ас студиед бy хуман браин потентиалс”. Ј Еxп Псyцхол. 21: 127—44. дои:10.1037/х0057431.
- ^ Демент, Wиллиам; Клеитман, Натханиел (1957). „Цyцлиц вариатионс ин ЕЕГ дуринг слееп анд тхеир релатион то еyе мовементс, бодy мотилитy анд дреаминг”. Елецтроенцепхалогр Цлин Неуропхyсиол. 9 (4): 673—90. ПМИД 13480240. дои:10.1016/0013-4694(57)90088-3.
- ^ Рецхтсцхаффен А, Калес А, едиторс. А Мануал оф Стандардизед Терминологy, Тецхниqуес анд Сцоринг Сyстем фор Слееп Стагес оф Хуман Субјецтс. Wасхингтон: Публиц Хеалтх Сервице, УС Говернмент Принтинг Оффице; 1968.
- ^ Ибер, C; Анцоли-Исраел, С; et al. (2007). The AASM Manual for the Scoring of Sleep and Associated Events: Rules, Terminology and Technical Specifications. Westchester: American Academy of Sleep Medicine.
- ^ World, Psychology (1998). „Stages of Sleep” (PDF). Архивирано из оригинала (PDF) 26. 06. 2008. г. Приступљено 15. 6. 2008. „(includes illustrations of "sleep spindles" and "K-complexes")”
- ^ Schulz, H. (april 2008). „Rethinking sleep analysis”. Journal of Clinical Sleep Medicine. 4 (2): 99—103. PMC 2335403
 . PMID 18468306.
. PMID 18468306.
- ^ Cirelli, Chiara; Tononi, Giulio (26. 8. 2008). „Is Sleep Essential?”. PLoS Biol. Public Library of Science. 6 (8): e216. PMC 2525690 . PMID 18752355. doi:10.1371/journal.pbio.0060216. Архивирано из оригинала (Essay) 11. 09. 2008. г. Приступљено 21. 4. 2009. „... it would seem that searching for a core function of sleep, particularly at the cellular level, remains a worthwhile exercise”
- ^ Daan S, Barnes BM, Strijkstra AM (1991). „Warming up for sleep? Ground squirrels sleep during arousals from hibernation”. Neurosci. Lett. 128 (2): 581. PMID 1945046. doi:10.1016/0304-3940(91)90276-Y.
- ^ а б в г д ђ е ж з и ј Sadock, B., Alcott Sadock, V. (2005). Kaplan & Sadock's Comprehensive Textbook of Psychiatry (Eighth Edition). Philadelphia: Lippincott Williams & Wilkins.
- ^ National Geographic Magazine, May 2010, "The Secrets of Sleep", by D.T. Max.
- ^ Dement, Wiliam (10 June, 1960). „The Effect of Dream Deprivation”. American Association for the Advancement of Science. Архивирано из оригинала 28. 04. 2024. г. Приступљено 18 September, 2016. Проверите вредност парамет(а)ра за датум:
|access-date=, |date=(помоћ) - ^ Kryger, M., Roth, T., Dement, W. (2011). Principles and Practise of Sleep Medicine (Fifth Edition). St. Louis: Elsevier Saunders.
- ^ Sleep Deprivation, Psychosis and Mental Efficiency Архивирано на сајту Wayback Machine (29. август 2008), Станлеy Цорен Пх.D., Марцх 1998, Вол. XV, Иссуе 3
- ^ а б http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1781306/
- ^ ПРНП-гене
- ^ "Тхе Сецретс оф Слееп", бy D. Т. Маx, Натионал Геограпхиц Магазине, Маy (2010). стр. 74.
- ^ Турнер, Ребецца. „Тхе Ман Wхо Невер Слепт: Мицхаел Цорке”. Wорлд Оф Луцид Дреаминг. Приступљено 20. 5. 2011.
- ^ Марић, Ј. (2005). Клиничка психијатрија (једанаесто, прерађено и допуњено издање). Београд: Меграф.
- ^ „Гене Цутс Неед фор Слееп - Слееп Дисордерс Инцлудинг, Слееп Апнеа, Нарцолепсy, Инсомниа, Сноринг анд Нигхтмарес он МедицинеНет.цом”. Архивирано из оригинала 14. 07. 2011. г. Приступљено 11. 6. 2010.
- ^ Хе, Y.; Јонес ЦР; Фујики, Н.; et al. (август 2009). „Тхе трансцриптионал репрессор ДЕЦ2 регулатес слееп ленгтх ин маммалс”. Сциенце. 325 (5942): 866—70. ПМЦ 2884988 . ПМИД 19679812. дои:10.1126/сциенце.1174443.
- ^ Хрњица, С. (2005). Општа психологија са психологијом личности (десето издање). Београд: Научна књига нова.
- ^ а б в г Пурвес, D. ет ал. (2008). Неуросциенце (Фоуртх Едитион). Сундерланд: Синауер Ассоциатес, Инц.
- ^ Пурвес, D., Аугустине, Г., Фитзпатрицк, D., Халл, W., ЛаМантиа, АС., МцНамара, Ј., & Wхите, L. (2008). Неуросциенце (Фоуртх Едитион). Сундерланд: Синауер Ассоциатес, Инц.
- ^ Мукхаметова, ЛМ; Супина, АY; Полyаковаа, ИГ (14. 10. 1977). „Интерхемиспхериц асyмметрy оф тхе елецтроенцепхалограпхиц слееп паттернс ин долпхинс”. Браин Ресеарцх. 134 (3): 581—584. ПМИД 902119. дои:10.1016/0006-8993(77)90835-6.
- ^ Фарацо, Јулиетте (1. 8. 2000). „Ре: Аре тхере анималс wхо дон'т слееп ор тхат слееп верy литтле?”. МадСци Нетwорк: Зоологy. Приступљено 25. 1. 2008.
- ^ ЛивеСциенце.цом, Инсомниа Маниа: Неwборн Маммалс Дон'т Слееп фор а Монтх.
Спољашње везе
уреди- (језик: енглески) Америчка академија за медицину спавања